
 CC-BY 4.0
CC-BY 4.0
Coral reefs are highly biodiverse and productive ecosystems (Reaka-Kudla, 1997; Fisher et al., 2015) that substantially contribute to human well-being (Moberg & Folke, 1999; Adey, 2000). Yet, reef-building corals are imperilled by rising temperatures and other anthropogenic stressors worldwide (Hughes et al., 2017; Knowlton et al., 2021; Souter et al., 2021). Thus, there is great urgency to inventory coral biodiversity and to deepen knowledge of biodiversity generating processes to guide conservation and restoration actions.
For corals, it has long been recognised that morphological variation is unlikely to align well with genetic delineation of biological units (Knowlton, 1993; Oury et al., 2023). Supporting this notion, Grupstra et al. (2024) found over 100 examples across 24 genera of genetic studies that self-reported discovering morphologically cryptic coral species. There is also evidence that many coral taxa have incomplete reproductive barriers and are connected by occasional gene flow or hybridisation (reviewed by van Oppen & Gates (2006); Willis et al. (2006); Mao & Satoh (2019); González et al. (2021); Hobbs et al. (2021); Pinsky et al. (2023)). Consistent with hybridisation, morphologically intermediate individuals between taxonomically-recognised species are often encountered in the field (Veron, 1995; Richards et al., 2008; DeVantier et al., 2020), and many species can be crossed under experimental conditions (Isomura et al., 2016; Chan et al., 2019; Kitanobo et al., 2022). Thus, there is a growing sense that natural units of coral biodiversity may often consist of genetically distinct groups that can be connected by gene flow (hybridisation), even if these groups are not necessarily morphologically distinct. Figure 1 shows examples of such cryptic genetic groups.
Documenting the prevalence of cryptic genetic groups among corals is an important first step towards learning how differentiated groups may be locally adapted to different environments and understanding the conditions under which their differentiation leads to speciation, fusion, or some in-between state of differentiation-with-gene flow (Abbott et al., 2013; Barraclough, 2024). Scenarios where divergence between groups has been evolutionarily recent and/or accompanied by gene flow present the greatest challenge for recognising distinct groups and are therefore underpowered to recognise mildly differentiated genetic groups, much less to discern the processes contributing to their coexistence when inferences rely on few loci (Seehausen et al., 2014). Thus, in this review, we focus exclusively on population genomic studies (supported by whole genome sequencing or reduced representation sequencing of genome-wide variation) and apply clear and reproducible criteria for recognising distinct genetic groups aligned to speciation theory and concepts. This paper builds upon important earlier reviews and syntheses of cryptic coral diversity (Richards et al., 2016; Grupstra et al., 2024) and hybridisation in corals (van Oppen & Gates, 2006; Willis et al., 2006; Hobbs et al., 2021; Pinsky et al., 2023) that primarily focused on studies with few markers (microsatellites, few sequenced loci, or allozymes).
We use the term cryptic to signify cases where there are distinct genetic groups among sets of colonies that were field-identified as being the same species by experienced researchers (consistent with the definition of Grupstra et al. (2024)). Thus, this definition may include both pseudo-cryptic genetic groups and those that truly lack morphological differences (Pante et al., 2015; Cahill et al., 2024) such that sensu lato would be the most appropriate epithet (Chenuil et al., 2019). These genetically distinct coral groups may not have all the contingent properties of species (de Queiroz, 2005), including complete reproductive isolation (as required by the biological species concept: Mayr, 1942). (Indeed, it is possible that some distinct genetic groups represent ecotypes, or genetically distinct populations adapted to specific environments, where their speciation outcome is uncertain: Lowry, 2012.) In this review, we use the terms genetic groups or genetic taxa to emphasize the genetic coherence (Dobzhansky, 1937) and distinctiveness (following Mallet’s (1995) operational definition) of genetically defined coral groups, yet acknowledge ambiguity in whether these groups should be taxonomically defined as species (or subspecies). We avoid terms such as clade or lineage, since monophyly across all or many gene trees is unlikely when divergence is recent.

Figure 1 – Examples of closely related taxa that are genetically differentiated, yet morphologically cryptic (sensu lato). Details can be found in the original studies: Stylophora pistillata (Meziere et al., 2024), Porites sp. (Starko et al., 2023), and Agaricia lamarcki (Prata et al., 2022).
It is likely that many cryptic coral taxa inhabit the “grey zone” of the speciation continuum, where taxa are linked by continuing gene flow that may be variable in strength across the genome (Seehausen et al., 2014; Roux et al., 2016; De Jode et al., 2023), and where taxonomy is likely to be controversial (de Queiroz, 2005; Roux et al., 2016). When reproductive barriers are not complete, the outcome of interbreeding between groups is controlled by the relative strength of divergent selection (when different alleles are advantageous for different groups) and the genomic extent of gene exchange (which promotes homogenisation) (Abbott et al., 2013). However, for genetically distinct groups to coexist within their dispersal range and across time, reproductive barriers of some form must be present (Coyne & Orr, 2004; Seehausen et al., 2014). For corals, reproductive barriers between groups may be due to intrinsic genetic incompatibilities (Levitan et al., 2004), differences in spawning times (Levitan et al., 2004; Rosser, 2015), and strong divergent selection arising from microhabitat differences (Prada & Hellberg, 2013).
Here, we critically assess the prevalence of cryptic genetic taxa among corals. By corals, we refer to benthic Anthozoans including scleractinians (hard corals) as well as octocorals (soft corals and gorgonians). In contrast to previous reviews on corals, we only consider population genomic surveys that have the power to detect cryptic taxa, and we reexamine reported results against reproducible and conservative criteria for detecting cryptic taxa based on population genetic evidence for some degree of reproductive isolation (in the spirit of Mayr’s 1942 biological species concept and related concepts stressing evolutionary lineages and populations: de Queiroz, 1998; see section 2.1 for more details). Specifically, we: 1) rigorously quantify the prevalence of cryptic coral taxa and hybridisation, 2) discuss the relevance of cryptic coral taxa and hybridisation for conservation and restoration, and 3) highlight new directions to develop corals as exciting model systems for speciation and adaptation studies that can bring valuable insights to conservation and restoration. The main text is supported by text boxes that (i) provide a worked example of delineating coral taxa, (ii) outline best practices for designing studies when cryptic taxa are likely to be encountered, and (iii) demonstrate how cryptic taxa are commonly overlooked in coral experiments.
To gauge the prevalence and impacts of cryptic coral taxa and hybridisation, we undertook a structured literature search and focus exclusively on population genomic studies of corals. Population genomic surveys have the power to detect subtly differentiated genetic groups when allele frequency differences between groups are small, as we expect for groups that are recently diverged and/or connected by gene flow (Section 2.1). Below, we estimate the frequency of cryptic genetic taxa (2.2). We describe what population genomic results have shown regarding the relationships between cryptic coral taxa and their symbiont partners (2.3) and the environment (2.4). We then investigate how often studies test for and observe hybridisation and gene flow between cryptic taxa (2.5).
2.1 Criteria to detect and delineate cryptic coral taxa
We define cryptic coral taxa as distinct groups of individuals (genotypic clusters, in line with Mallet, 1995) within nominal species that maintain their distinctiveness even when their ranges overlap, and therefore there are no physical barriers to gene exchange. Instances where distinct genetic groups are found together (i.e., sympatric within the scale of dispersal distance) provide the strongest circumstantial evidence for some degree of reproductive isolation between groups, as reduced gene flow due to restricted dispersal cannot be the primary cause of genetic divergence in these cases (Coyne & Orr, 2004; Seehausen et al., 2014). When distinct genetic groups are geographically separated (i.e., allopatric), then divergence may solely reflect physical dispersal barriers to gene exchange and therefore distinct allopatric groups are not informative for inferring reproductive isolation.
The statistical power of common methods for detecting population genetic structure is determined by the number of loci examined, as well as the extent of genetic covariance among loci, where distinct populations exhibit non-random associations of alleles at various loci across the genome (i.e., linkage disequilibria) (Pritchard et al., 2000; Novembre et al., 2008). While physical linkage on a chromosome alone will cause covariance among loci, genome-wide covariance also arises as a direct consequence of population structure (reflecting distinct gene pools subject to independent outcomes of genetic drift and selection) or selection on interacting loci. Reproductively isolated taxa and geographically separated populations will be differentiated from each other across loci due to genetic drift and selection. In contrast, gene flow between taxa will reduce allele frequency differences and covariances. Statistical power for identifying small allele frequency differences and linkage disequilibria in empirical surveys is increased by sampling many individuals (both within and between locations) and many loci, where important genomic differences between groups can be missed when few individuals or few loci are sampled. For example, two co-occurring genetic groups of Montastraea cavernosa were clearly delineated using thousands of loci and yet were ambiguous with 9 microsatellite loci (Sturm et al., 2020). Similarly, chromosomal inversions have been found in Acropora kentii using whole genome sequencing (Zhang et al., 2024) that were missed with single nucleotide polymorphisms from a reduced portion of the genome (Matias et al., 2023).
After adequate sampling and with sufficient genotyped loci, there are two general approaches used for detecting subtle genome-wide differentiation: ordination-based analyses and model-based clustering. Ordination-based analyses, such as principal components analyses (PCA), principal coordinates analysis, and multidimensional scaling describe multidimensional relationships among entities and are based on (and/or visually represent) the genetic covariance matrix (Patterson et al., 2006). Model-based clustering analyses – typified by admixture detection analyses such as STRUCTURE (Pritchard et al., 2000), fastSTRUCTURE (Raj et al., 2014), ADMIXTURE (Alexander et al., 2009) and sNMF (Frichot et al., 2014) – partition groups (K) based on associations among alleles and loci. Ordination and model-based clustering approaches are valuable for exploring relationships between individuals without pre-assigning individuals to “populations”, as is required by F-statistics and other population-level metrics. These methods are some of the most common and routinely employed methods in population genomic surveys, perform well at low levels of divergence (i.e. where allele sharing is prevalent), and provide complementary insights into spatial patterns of genetic diversity. We omit results from discriminant analysis that maximise variance using user-assigned groupings (see Thia (2022) for extended discussion).
Specifically, we propose three criteria for identifying and delineating taxa based on ordination and model-based clustering:
-
Distinct genetic groups occur in sympatry relative to their dispersal ability.
-
Ordination analyses (e.g. PCA) strongly cluster these distinct genetic groups based on genotypes of individuals and/or model-based clustering indicates that individuals belong to separate groups.
-
Genetic distances between sympatric individuals of provisionally different taxa are greater than the genetic distances among allopatric individuals (when allopatric individuals comprise a single putative taxon). This is evidenced by divergence between sympatric groups across lower ordination axes and/or group numbers (K values) for clustering as compared to axes or groups that describe geographic structure.
2.2 Evaluating published studies for evidence of cryptic taxa
To determine if published genomic surveys of corals typically test for and find evidence of cryptic taxa following the above criteria (2.1, and see Box 1 for worked example), we searched the Web of Science Core Collection database for published papers displaying graphical results from ordination (PCA, primarily) and/or model-based clustering analyses (STRUCTURE, fastSTRUCTURE, ADMIXTURE, and sNMF). We focus on studies that genotyped individual cnidarian genomes for 1000s of loci or more and sampled across two or more geographic locations (see Appendix for details of literature search). We additionally collected information on depth ranges and symbiont composition when reported. In our review of published studies, we apply our three criteria (from Section 2.1) conservatively by only looking for sympatric differentiation: i) along the first principal component axis (referred to as ‘PC1’) for ordination-based analyses, and ii) for model-based clustering, examining outcomes when individuals were allowed to be assigned to one of two groups (i.e. K=2). We consider individuals sampled ≤10 km apart as being broadly sympatric. Dispersal distances for most coral species are unknown but there is evidence of spatial population structure at scales of 10 km in brooders (Prata et al., 2024), which guided our choice for sympatric distance.520
Box 1: Applying taxonomic delineation with reproductive isolation criteria
We propose three requirements for identifying and delineating coral taxa using genomics-informed ordination and model-based clustering approaches (Section 2.1). In the empirical example that follows, based on Prata et al. (2024, with methods detailed in their supplementary files), we outline how coral cryptic taxa were identified using these three criteria and highlight difficulties with their interpretation.
Colonies of the brooding coral, Agaricia agaricites, were sampled at four locations ~10-15 km apart along west Curaçao, and collections were further subset into three depths (5, 10 and 20 m) at each location. Genotyping using reduced representation sequencing of 335 colonies and 1,629 SNP-loci revealed distinct genetic groups co-occurring within four sampled sites. This study provides a clear example of cryptic taxa identified according to criteria 1-3. Furthermore, taxa occupied unique depth ranges (AA1 occurs predominantly at 20 m, whereas AA2 occurred at all depths sampled) suggesting divergence of taxa by habitat (Fig. T1A & C).
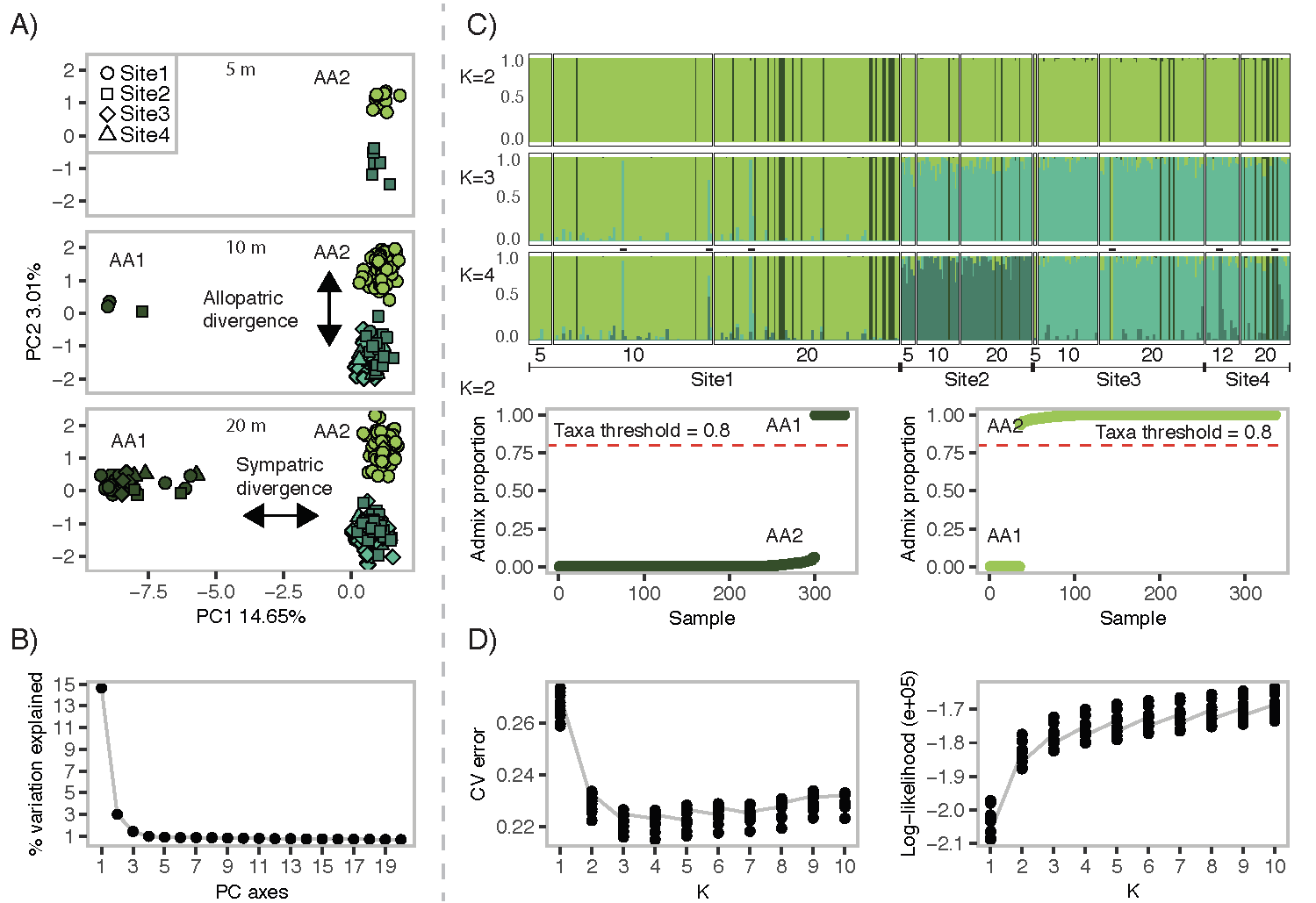
Figure T1 - Agaricia agaricites resolved into two distinct taxa and fulfil criteria 1, 2 and 3 for cryptic taxon delineation. A) PC1 resolves two sympatric groups at every sampling site. PC2 represents an example of geographic partitioning in AA2 (Site 1 vs Site 2 - 4) and therefore does not necessarily imply intrinsic reproductive isolation. B) Shows that the percentage of variation explaining PC1 (14.65%) is ~5x more than PC2 (3.01%). C) Analyses using ADMIXTURE for K=2 assigned individuals to each group with high confidence (q > 0.9) and showed that AA1 and AA2 were sympatric at all sites. D) There is a significant drop in cross-validation error between K=1 and K=2, and greater log-likelihood, supporting the selection of K = 2. All three criteria are met in delineating AA1 versus AA2 as cryptic taxa within A. agaricites.
This example also illustrates one potential complication for interpreting differentiation among putative taxa – the presence of recent migrants. The second PC axis (also mirrored in ADMIXTURE results for K=3 and K=4) shows partitioning that largely aligns with geographic separation and would not be considered as delineating distinct taxa under our criterion of sympatry (criterion 1 in 2.1). If variation captured by the second PC axis (and K=3 and K=4) reflects geographic differentiation, then geographically mismatched genotypes likely reflect recent immigration (shown by dashes among K=3 and K=4 in Fig. T1C). Distinguishing migrants from distinct taxa may be especially difficult when sampling numbers are low. However, if gene flow is high (> 1 migrant per generation) and there are no barriers to reproduction, then the structure between populations is expected to dissipate over a few generations (Waples & Gaggiotti, 2006).
While this example highlights the utility of PCA and cluster-based modelling methods for identifying cryptic coral taxa, patterns shown in these analyses are not always straight-forward to interpret. Additional subsetting and filtering steps are necessary to thoroughly scrutinise data for consistent patterns and reveal accurate groupings (see Supplementary in Prata et al. (2024)). To better understand the possible biases of both PCA and assignment methods, we refer readers to McVean (2009), Pritchard et al. (2000), and Puechmaille (2016). More detailed discussions on species delineations, especially for marine species, can be found in Pante et al. (2015). Once taxa are delineated, investigators can investigate signals of recent hybridisation (e.g. Anderson & Thompson (2002)) and test among demographic models of historical gene flow and divergence (as in Roux et al. (2016) and De Jode et al. (2023)) to better understand the nature of population divergence (see section 2.5).
The literature search uncovered 41 studies describing results for 31 species. Some studies included multiple species, and some species were genotyped multiple times in different studies, thus, our search yielded a total of 51 unique records (available as supplemental data). Although we did not restrict our search by sampling depth, none of the recovered records included species beyond mesophotic depths (i.e., >150 m), and therefore the results that follow describe shallow water and mesophotic corals.
As shown in Fig. 2, 68% of nominal species with population genomic data showed evidence for distinct cryptic taxa (representing 23 records out of 39 that could be evaluated against all three criteria). While we might expect genetic differentiation to be greater among brooding corals that have less innate dispersal ability relative to broadcast spawning corals (Knowlton, 2001), the relative proportions of sympatric versus non-sympatric groups did not differ between brooding and broadcast spawning corals (2x2 Fisher’s exact test using either ordination or clustering, P > 0.2).
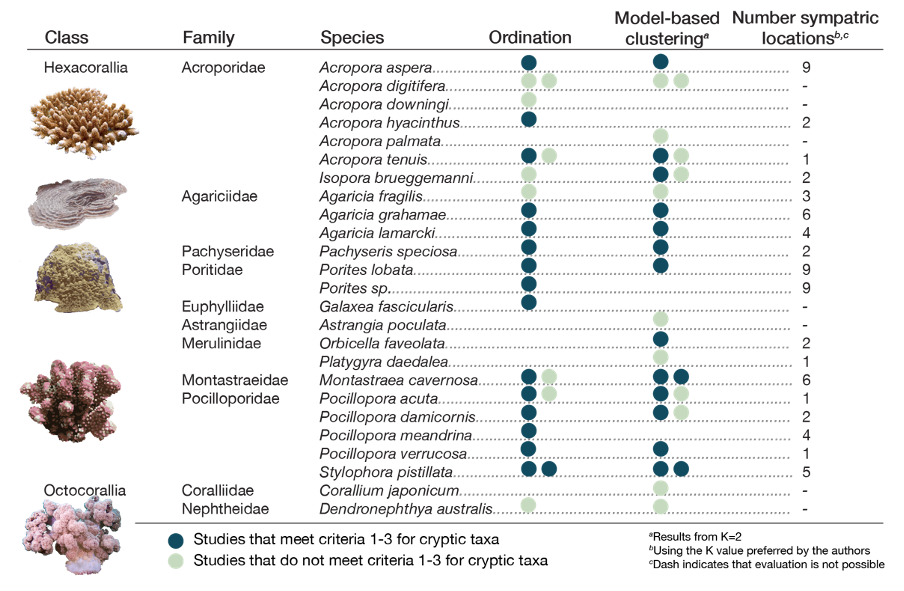
Figure 2 – Evidence for cryptic coral taxa is common. For many genomic studies of corals, the greatest axis of genetic differentiation defines groups that are sympatric (e.g. satisfying criteria 1-3 for cryptic taxa). Results by species are summarised as either meeting or not meeting the three criteria for cryptic genetic taxa as applied to ordination or model-based clustering results. For criterion 3, we apply the strictest definition where sympatric differentiation is aligned to the first axis (ordination) or K = 2 (model-based clustering). From the 51 studies examined, 39 presented results that could be evaluated against criteria 1-3. Some studies had both ordination and model-based clustering, and some species were included in more than one study; thus, multiple points can appear against each species. Of the 25 nominal species with population genomic data, 17 showed evidence for including cryptic taxa.
Criterion 3 is based on genetically distinct groups co-occurring. This evidence is stronger when these groups co-occur across many geographic locations. Thus, we investigated whether pairs of genetically distinct coral groups were repeated across multiple sites using model-based clustering results based on author-selected K values. Focusing on reported results from principal components analyses and model-based clustering allowed us to evaluate patterns across a broad cross-section of published studies. Ideally, the processes responsible for creating such patterns should be further evaluated, where demographic inference can be used to estimate gene flow and thereby provide greater insights on reproductive isolation (see for example, Fraïsse et al. (2021) and section 2.5). Across studies, it was common for cryptic groups to be sympatric at multiple sampled sites (Fig. 2). This observation strengthens the conclusion that closely related, but distinct, taxa can co-occur over extensive geographic areas and implicates some degree of reproductive isolation maintaining the distinctiveness of each group (discussed further in 2.5). In summary, cryptic taxa are common attributes in population genomic studies of corals.
Although there is evidence for cryptic taxa across many studies (Fig. 2), not all studies acknowledged the groupings within their data or partitioned their data appropriately in downstream analyses. For example, some of the studies computed summary statistics such as heterozygosity and F-statistics using all individuals from the sampling locations, despite evidence for genetically distinct taxa co-occurring within locations and thereby created inaccurate estimates (discussed further in 3.3). Indeed, many previously published studies that did not have the advantage of detecting cryptic taxa reliably with genomic data (e.g. inference based on microsatellites or allozymes) have likely inadvertently based conclusions on heterogeneous mixes of cryptic taxa. Additionally, analyses using few loci likely lack sufficient power to detect recently differentiated taxa, and thus underpowered studies may conclude the absence of cryptic taxa without sufficient evidence. For these reasons, many published studies – including studies published by authors of this review – may unintentionally base conclusions on heterogeneous mixes of cryptic taxa.
2.3 No clear patterns for symbionts associated with cryptic coral host taxa
An important aspect of coral biodiversity is the diverse microbial community living within the cnidarian host, where mutualistic relationships with endosymbiotic dinoflagellates from the family Symbiodiniaceae are known to affect whole organism physiology (LaJeunesse et al., 2018). A central issue for coral diversification and adaptation is the extent to which symbiotic associations are flexible (see Grupstra et al. (2024) for an extended discussion). We found that 43% of surveyed genomic studies had genotyped dinoflagellate symbionts alongside the coral hosts. Symbiont strains were commonly found to be shared across cryptic host taxa regardless of reproductive mode (Bongaerts et al., 2017; van Oppen et al., 2018; Gomez-Corrales & Prada, 2020; Bongaerts et al., 2021a; Prata et al., 2022; Rivera et al., 2022; Buitrago-Lopez et al., 2023; Starko et al., 2023). Yet, some symbiont strains, appeared specific to cryptic taxa (van Oppen et al., 2018; Gomez-Corrales & Prada, 2020; Bongaerts et al., 2021a; Buitrago-Lopez et al., 2023; Starko et al., 2023) so there was no single consistent pattern relating symbionts with their coral hosts, as is commonly observed across nominal species (Bongaerts et al., 2015). However, the variety of methods used to characterise symbionts may contribute to inconsistent patterns.
Moving beyond single marker genotyping of symbionts may provide better resolution of host-symbiont associations (Davies et al., 2023; Ishida et al., 2023; Zhang et al., 2023). For example, Rivera et al. (2022) found that symbiont identities among Porites lobata did not align to host taxa using internal transcribed spacer (ITS) genotyping, but instead were concordant with higher resolution SNP-based analyses. Among the studies examined here, many (e.g. Howells et al. (2016); van Oppen et al. (2018); Gomez-Corrales & Prada (2020); Bongaerts et al. (2021a); Prata et al. (2022); Buitrago-Lopez et al. (2023); Starko et al. (2023)) relied on ITS sequences to characterise within-colony symbiont lineages. Some studies used incidentally recorded symbiont sequences retrieved from whole-colony sequencing (either reduced representation or shotgun whole genome sequencing) to make inferences about symbionts, including reconstructing symbiont organelle diversity (Bongaerts et al., 2017; Forsman et al., 2017; Gonzalez-Zapata et al., 2018; Cooke et al., 2020; Bongaerts et al., 2021a; Matias et al., 2023; Zhang et al., 2023) or characterising symbiont genomic diversity with k-mer analyses (Zhang et al., 2023). Of these approaches, k-mer analysis is the only method that captures genome wide diversity of symbionts and therefore may reveal more nuanced patterns than ITS or organelle-based results (which rely on a single marker) (Ishida et al., 2024).
2.4 Depth can segregate cryptic genetic taxa
For coral hosts, the existence of distinct genetic groups in sympatry implies that differentiation could be preserved by strong divergent selection arising from adaptation to local environments (Richardson et al., 2014). Such divergent selection could maintain ecotypes within species and, in extreme cases, drive reproductive isolation (ecological speciation: Schluter (2001); Rundle & Nosil (2005)). It has been hypothesised that ecological speciation in corals could be common, resulting from strong environmental gradients on reefs, especially associated with depth (González et al., 2018). Indeed, depth is a predominant structuring aspect in coral reef communities (Knowlton, 1993), with distinct species turnover between shallow (approximately < 30 m) and mesophotic (approximately 30-150 m) depth zones. Importantly, these transitions exist across communities because many environmental factors covary with depth, such as light intensity and spectrum (Lesser et al., 2009), temperature (Kahng et al., 2019), nutrients (Leichter et al., 2008), water flow (Muir et al., 2015), as well as disturbance frequency and severity (Bongaerts & Smith, 2019), creating highly contrasted habitats often only metres apart. Thus, selective agents could be both strong and multifarious and therefore depth is likely to partition taxa into ecotypes, matching patterns of species turnover.
Among the population genomic studies we surveyed, 11 studies undertook sampling over replicated depth-associated habitat contrasts (i.e., more than one site with two depth habitats sampled within each site). Replicated differentiation by depth was found for Agaricia fragilis (shallow vs. mesophotic, Bongaerts et al. (2017)), Agaricia lamarcki (shallow vs. mesophotic, Prata et al. (2022)), Isopora brueggemanni (lagoon vs. slope; Thomas et al. (2019)), Pocillopora damicornis (flat vs. slope, van Oppen et al. (2018)) and Montastraea cavernosa (shallow vs. mesophotic, Sturm et al. (2022)) but not for Agaricia grahamae (upper vs. lower mesophotic, Prata et al. (2022)), Stephanocoenia intersepta (i.e., shallow vs. mesophotic, Bongaerts et al. (2017)), Acropora digitifera (i.e., lagoon vs. slope, Thomas et al. (2019)), or Agaricia undata (shallow vs. upper mesophotic and upper vs. lower mesophotic, Gonzalez-Zapata et al. (2018)). Thus, differentiation by depth frequently, but not always, discriminated cryptic coral taxa as identified by our criteria (see Grupstra et al. (2024) for further discussion and examples).
The structure and composition of coral-associated microbial communities also can vary along environmental gradients, including those associated with depth and disturbance (Klaus et al., 2007; Bongaerts et al., 2013; Howells et al., 2013; Quigley et al., 2022). Shifting compositions of coral-associated microbial communities can expand the environmental niche available to the coral holobiont, mediating adaptation to environmental stress. Indeed, several of the studies examined here reported greater spatial or environmental partitioning among symbionts as compared to hosts (e.g. Astrangia poculata: Aichelman & Barshis (2020); P. verrucosa: Buitrago-Lopez et al. (2023); Stylophora pistillata: Buitrago-Lopez et al. (2023); Platygyra daedalea: Howells et al. (2016); Acropora tenuis: Matias et al. (2023); A. lamarcki: Prata et al. (2022); but not so for A. digitifera: Zhang et al. (2023)). Intriguingly, Starko et al. (2023) demonstrated that a distinct symbiont community associated with one cryptic taxon of massive Porites sp. shifted following a heatwave, such that the post-heatwave composition better matched the symbiont communities living in the other two cryptic taxa. Thus, symbiont communities may shift to track local environments (Baker, 2003), although this flexibility is likely to differ among host taxa (Quigley et al., 2022). How environmentally induced selection shapes genetic and phenotypic variation across biological partners of the coral holobiont remains an open question and would likely benefit from manipulative experiments.
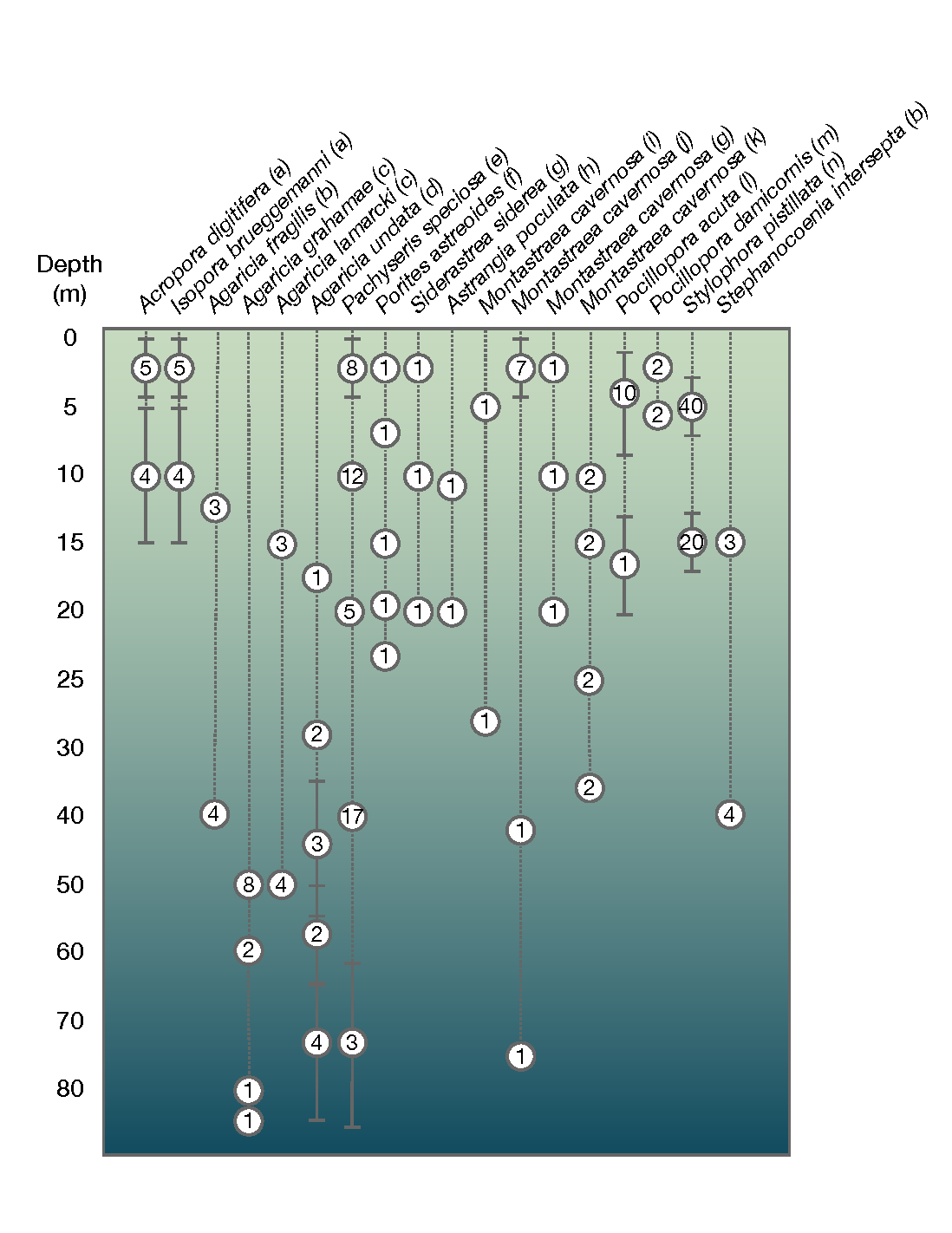
Figure 3 - Summary of depth sampling schemes for studies that reported depth and sampled at more than one depth. Numbers indicate the number of distinct locations that were sampled per depth. Locations were considered distinct if the nearest locations were depth contrasts (e.g. adjacent sites sampled at 5 and 15 m were considered as two locations); otherwise, locations within 10 km and at the same depth were collapsed to a single point. Dotted lines connect locations from the same study and thick grey lines indicate the sampling range (as reported by authors). Citations are as follows: a) Thomas et al., 2019; b) Bongaerts et al., 2017; c) Prata et al., 2022; d) Gonzalez-Zapata et al., 2018; e) Bongaerts et al., 2021a; f) Shilling et al., 2023; g) Rippe et al., 2021; h) Aichelman & Barshis, 2020; i) Drury et al., 2020; j) Sturm et al., 2020; k) Sturm et al., 2022; l) Aurelle et al., 2022; m) van Oppen et al., 2018; n) Meziere et al., 2024.
Whereas differentiation by depth and habitat appear to be common in corals (and their associated microbes), sampling strategies for many coral genomic studies are not designed to detect genetic differentiation along these environmental variables. Among the population genomic studies examined here, 25% failed to report sampling depth (or any other relevant habitat, including our own work, e.g. Matias et al. (2023)). Presumably, most of the genotyped corals across the studies we reviewed were collected on SCUBA from < 30 m and from a similar depth range across all sites. Among studies that did report depth, many sampling regimes had depth confounded with geography, where each location was sampled at a single depth (Fig. 3). A minority of studies (21%) implemented a structured sampling design where the same depth was sampled at more than one location. More complete reporting on depth and other microenvironmental attributes alongside sampling study designs that replicate environmental contrasts are needed to advance our understanding of how heterogeneous environments, divergent selection, and intrinsic reproductive isolation interact to shape coral biodiversity.
2.5 Gene flow links coral taxa across divergence histories
Having established the prevalence of cryptic coral taxa (2.2), and their microbial (2.3), and abiotic (2.4) associations, we re-examine coral population genomic studies for evidence of gene flow and contemporary hybridisation between cryptic taxa. Gene flow between coral taxa has long been suspected (Veron, 1995; van Oppen & Gates, 2006; Willis et al., 2006), but studies using few genetic markers often lack the resolution to appropriately investigate hybridisation in the context of recently diverged taxa, where genetic similarities can result from either shared ancestral diversity or gene flow (discussed in 2.1). Thus, by analysing thousands of genomic SNPs, genomic studies can often resolve the likelihood of divergence with gene flow versus strict divergence scenarios, identify genomic regions that have experienced high or low levels of gene flow, and find regions where introgression patterns have been shaped by selection (Taylor & Larson, 2019).
To gauge if cryptic coral taxa have been linked by gene flow over their divergence history, we focused on the few coral population genomic studies from our literature search that undertook demographic modelling of speciation histories between cryptic genetic groups. Population genetic demographic modelling involves comparing the probability of alternative historical scenarios (e.g. no gene flow versus periodic or ongoing gene flow: Gutenkunst et al., 2009; Beaumont, 2010; Sousa & Hey, 2013; Fraïsse et al., 2021) to resolve the relative contributions of shared ancestral polymorphisms and gene flow to shared genetic variation among taxa. Eight studies used demographic modelling to evaluate competing divergence scenarios between cryptic taxa (e.g. moments and δaδi methods, representing 11 nominal species: Fig. 4). Strikingly, all 12 evaluated records found the greatest support for models involving periods of divergence with gene flow. Among the variety of divergence with gene flow scenarios supported, only one record (Cooke et al., 2020) found exclusive support for the secondary contact model over other scenarios, while four records had the greatest support for divergence with continuous gene flow (isolation migration model). Other models either could not distinguish between scenarios or supported models that included periods of isolation after initial divergence with gene flow (Fig 4). In all these examples, gene flow was evolutionarily significant, but divergence was sufficient to overcome the homogenising effects of gene flow. Because divergence is maintained, gene flow between groups cannot be occurring at high enough rates to boost census population sizes, that is, it is not ecologically significant (Waples & Gaggiotti, 2006).
The frequent rejection of secondary contact models can be used as evidence for divergence without physical barriers to gene flow (Prada & Hellberg, 2021; De Jode et al., 2023), consistent with ecological speciation (Schluter, 2001; Rundle & Nosil, 2005). In practice, however, secondary contact and isolation migration models can be difficult to distinguish unless secondary contact is recent and is preceeded by a long period of isolation (Roux et al., 2016). Because all studies reported divergence times that predate Holocene reef configurations (< 10,000 years), the genetic groups observed today will have undoubtedly shifted range positions in response to past changes in coastal and reef configurations, and therefore, intermittent periods of isolation cannot be firmly ruled out in the models that support continuous gene flow (see similar arguments in De Jode et al., 2023 for speciation histories across diverse marine taxa). Periods of isolation facilitate the evolution of intrinsic reproductive barriers between groups (Seehausen et al., 2014), which can magnify genotype-by-environment associations such as differentiation by depth (Bierne et al., 2011). Thus, it will be important for future investigations to experimentally measure the strength and nature of reproductive barriers among cryptic coral groups to make inferences about genes associated with local adaptation (discussed further in section 4).

Figure 4 – Estimated divergence times of coral cryptic taxa vary from 100 thousand to millions of years but taxa are consistently connected by gene flow, as shown by demographic inferences across a variety of methods. Histories of divergence with gene flow were supported in all tested instances and similarly heterogenous gene flow across genomes was found in all tests (consistent with genomic islands of differentiation). Models follow conventional abbreviations: IM = isolation migration; SC = secondary contact; AM = ancient migration, PER = periodic migration, and SI = strict isolation. The only model that precludes gene flow throughout the entire divergence history is strict isolation. Models with an appended ‘a’ (e.g. ‘IMa’) signify a model with asymmetric migration, otherwise migration was modelled as symmetric. In brackets are alternative models that were tested and discarded. An asterisk in the bracket signifies that there was extensive model testing including the standard scenarios listed above. Dashes indicate information or parameters that were either not tested or not reported.
An emerging observation across a diversity of metazoans is that gene flow between closely-related species is variable across genomes (Ravinet et al., 2017) due to reproductive incompatibilities or local adaptations that reduce gene flow within some genomic regions (Martin & Jiggins, 2017; Ravinet et al., 2017). For example, chromosomal inversions and other genome features that lower recombination are thought to assist adaptive divergence when there is ongoing gene flow (Seehausen et al., 2014). However, whether these features maintain existing adaptive differentiation in corals is largely unexplored (Zhang et al., 2024). For pairs of sympatric coral taxa, demographic models that included heterogeneous gene flow rates received the highest support (Fig. 4). These results provide indirect evidence for genomic regions that are resistant to gene flow in the sympatric taxa studied, despite gene flow affecting neutral parts of the genome. For example, in S. pistillata, more divergent taxa had a higher proportion of their genomes experiencing reduced gene flow compared to the less divergent taxa, implying that genomic islands of differentiation become wider as speciation proceeds (Meziere et al., 2024). These findings are consistent with S. pistillata comprising morphologically similar taxa at various stages of divergence (Roux et al., 2016).
Low levels of gene flow can directly contribute to adaptation via introgression (Martin & Jiggins, 2017; Barraclough, 2024), where alleles derived from a different species can introduce adaptive traits into the receiving species (e.g. resistance to hypoxia at high altitude in humans: Huerta-Sánchez et al. (2015); and winter coat colour in hares: Giska et al. (2019)). In corals, a genomic region of approximately 220 kb appears to contribute to increased bleaching-tolerance for one Acropora hyacinthus taxon relative to other cryptic A. hyacinthus taxon, which may have been acquired through past hybridisation with Acropora millepora (Rose et al., 2021). Supporting this conclusion, there is evidence of historical Acropora range expansions coinciding with introgression events, suggesting that ecological opportunities and interspecies competition during range expansions contributed to Acropora diversification (Mao et al., 2018). While these studies implicate a role for hybridisation in adaptive evolution, there have been no comprehensive investigations of adaptive introgression in corals to date.
If interbreeding between divergent groups is ongoing and sufficiently common, then individuals with hybrid or mixed ancestry are likely to be present. Population genomic studies often find coral colonies with possible hybrid ancestries, but only two studies included explicit tests for recent hybridisation (such as tests implemented in NewHybrids: Anderson & Thompson (2002)) that probabilistically assign individuals as putative first-generation and early backcrosses. Early generation hybrid individuals were found among Agaricia taxa (Prata et al., 2022) but not among S. pistillata taxa (Meziere et al., 2024). Other studies have identified likely hybrid individuals based on the proportion of assignment to different groups from model-based clustering outputs (e.g. Cooke et al. (2020); Kitchen et al. (2020); Bongaerts et al. (2021a); Fifer et al. (2021); Rippe et al. (2021); Rivera et al. (2022); Matias et al. (2023)). Using a majority background group assignment score of <0.75 for distinct sympatric genetic groups as an indication of possible recent hybridisation, we identified potential hybrid individuals for 21 of the 34 species surveyed. These mixed ancestry individuals are based on the original author preferred K groups and therefore do not necessarily reflect admixture between partially reproductively isolated groups (see Box 1 for further discussion). Nonetheless, the prevalence of individuals with mixed ancestry suggests that hybridisation could be ongoing for many cryptic taxa. Notably, no population genomic study to date has tested whether putatively hybrid individuals were also morphologically or phenotypically distinct from parental individuals (Veron, 1995; Richards et al., 2008).
2.6 Summary: cryptic genetic taxa are common in corals
The preceding reanalysis and review of population genomic studies shows that cryptic coral taxa are common, estimated to be found in 68% of nominal species examined (Fig. 2, Section 2.2), and may be adapted to different microenvironments – especially depth (2.4). These cryptic taxa, however, are often linked by sharing symbiont strains (2.3 and 2.4) and via some gene exchange (2.5). Therefore, cryptic taxa may be distinct in terms of ecology, physiology, and evolution, but how to describe and delineate taxa is not clear, as there may not be obvious morphological characteristics to distinguish them. Even with access to genomic-scale genotyping, taxonomic resolution is affected by sampling and open to interpretation. It is clear, however, that gross morphology assessed by humans under field conditions is unreliable for recognising closely related taxa (Fig 1). Simply put, any coral investigation that does not genotype the corals under study risks treating a heterogeneous mix of partially reproductively isolated taxa as a single species. Box 2 suggests some pathways forward, where collaborations between geneticists and other coral reef scientists will play a key role.
Box 2: Best practice recommendations
Future surveys of corals should be designed with the expectation that cryptic species could be encountered. This means undertaking spatially and environmentally structured and replicated sampling, reassessing field collection protocols, and testing for cryptic taxa as part of bioinformatic and population genetic analyses. Ensuring that all data and metadata are thoroughly documented ensures future investigations can re-examine published data as novel analyses emerge, thus improving the re-use and re-purposing of genetic data.
Spatial sampling at the colony level
The best evidence for discriminating cryptic taxa from population structure is when distinct taxa are observed in sympatry (or close geographic proximity) at multiple locations (Section 2.1). We acknowledge that there is an element of chance in co-sampling distinct taxa. Given that depth appears to be the most common axis of differentiation, however, researchers who are planning to sample across depth should ensure that the same depths are sampled at multiple distinct locations to enable the detection of repeated co-occurrences of distinct taxa.
Alongside structured sampling, investigators would greatly enhance their data’s re-useability (and scope for future inference) by transitioning from a population sampling mindset to focusing on individual sampling and seeking to capture as much environmental context as possible at the colony level. For instance, “cryptic” species may in fact be morphologically distinguishable based on subtle characteristics (for example, S. pistillata, Meziere et al. (2024)) and therefore, could be considered “pseudo-cryptic”. Taking comprehensive photographs that can be examined later (see, for example, Protocol for Coral Collection & Curation by Project Phoenix: https://coralprojectphoenix.org/resources/#protocols) may allow diagnostic characters to be identified post hoc. An exciting avenue for new research could be to use photographs of genotyped cryptic taxa as training datasets for machine learning approaches to rapidly identify subtle differences in their micro-morphological characters (as expert taxonomists do already) to enable non-experts to identify pseudo-cryptic taxa in the field. For a subset of samples, it would be useful to retain larger colony fragments that would be suitable as museum voucher specimens (if permits allow) and maintain genetic vouchers for future studies. Recording each colony’s geoposition and depth can greatly support analyses based on depth (i.e. as a continuous rather than categorical predictor) and space, which simultaneously could provide insights on the microhabitat attributes of cryptic taxa and hybrid individuals (as in Prada & Hellberg (2014)). A particularly exciting technology that can greatly advance this colony-focused perspective is photogrammetry (Bongaerts et al., 2021b). We recognise that moving the focus from coral populations to colonies will require more time, effort, and greater expense, but the insights into potential cryptic taxa and coral biology more generally will be far richer.
Datasets that link genomic genotyping with ecological context at the colony level will be immensely valuable for gaining insights into ecological and evolutionary processes relevant to conservation. To maximise this value, investigators should strive to make all facets of their data FAIR compliant (Wilkinson et al., 2016), which includes linking genotypes with all recorded metadata, including metadata that might not be relevant to the original study (but that might be of value to other researchers, included with linked data files). Analytical pipelines also need to be fully reproducible by enabling consistency in bioinformatics and analytical decisions across studies such that outcomes can be confidently compared. No doubt, all this extra documentation is a substantial amount of work, and therefore should be forecasted in initial project planning. Coral biologists can take inspiration from plant population geneticists who have greatly advanced insights and impacts by sharing highly curated datasets that have been re-used to support a myriad of additional studies after their initial publication; for example, the IntraBioDiv (Meirmans et al., 2011) dataset of 27 co-distributed alpine plant genotypes has supported numerous reanalyses and test cases. Additionally, the genomic (and phenotypic) datasets for lodgepole pine and spruce from the AdapTree group (https://adaptree.forestry.ubc.ca/about/scientific-summary/) have greatly advanced our understanding of spatial adaptive diversity in trees.
Adjusting bioinformatic pipelines and analyses
Bioinformatics and population genetic analyses also need to be sensitive to the possibility of cryptic taxa (see also Section 2.1 and Box 1). Missing data thresholds and other data quality filters are employed as standard practice on individuals and loci. However, the sensitivity of different missing data thresholds to test taxon assignment and hierarchy hypotheses are often omitted. The more divergent groups are, the fewer sites they will share; thus, blanket missing data thresholds on heterogeneous samples may bias outcomes especially for sites with small sample sizes. Applying different missing data filters and subsetting datasets by selecting an even representation of predetermined groups (from initial model-based clustering analyses) or isolating certain groups can help in determining if the assignment and hierarchy of groups is stable and robust to the filters selected (Pritchard et al., 2000; McVean, 2009; Puechmaille, 2016). Intermediate or admixed individuals may appear as hybrid individuals, but the causes of these patterns are many, including unexplained variance due to geographic structure, under-sampled taxa, admixture with unsampled taxa, or higher levels of missing data for some individuals. Thus, we suggest formal hybrid tests be employed for clarification (e.g. NewHybrids) if datasets are suitable. Investigators should be transparent regarding how biases or decisions were handled when reporting groupings. We suggest following advice from Meirmans (2015) by always reporting multiple K values when using model-based clustering methods, as clustering analyses represent a heuristic approach that is open to interpretation for all biologically-sensible K values, even if an optimal K-value is selected by the user-defined summary statistic. Similarly, PCA results should present the percent of variation explained and include multiple axes (as there may be more than two cryptic groups and/or geographic structure within groups). Ultimately, we hope that the guidelines presented here can be used as a framework to detect coral cryptic taxa in future population genomic investigations.
When designing experiments
While population geneticists are the primary target audience for our recommendations, any coral biologist whose data interpretations could be affected by cryptic species would do well to incorporate genotyping in their project planning or minimally keep coral tissue for future genotyping. Our research group has found that preserving tissues quickly after collection (within 30 minutes post dive with samples placed in a cool container until processing), keeping tissue in 95% EtOH and replacing the liquid with fresh EtOH several times (until liquid is clear) within the first 48 hours of preservation, and ensuring there is a large EtOH to tissue volume ratio (≥10x) are critical steps for retaining high quality DNA. Other groups have had success with DMSO (Gaither et al., 2011) and ultra cold freezing (e.g. liquid nitrogen or -70 freezer) is always a good choice (although rarely feasible in field conditions). Pilot trials and consulting with geneticists who work on a particular species can point to appropriate methods. For experimental work, we propose that future studies should: 1) where possible, include larger sample sizes (n > 30) to screen for cryptic genetic population structure (this will ensure downstream comparisons in individual phenotypic differences are not confounded by cryptic speciation); 2) follow guidelines from 2.1 to recognise cryptic species; 3) report initial data checking methods and results (e.g. screening population structure) in publications and reports to assist the interpretation of individual- and population-level differences; and 4) clarify definitions and conventions for terms such as “cryptic species” and establish common terminology. If genotyping cannot be combined with the original study, then keeping preserved tissues and associated records will allow future genotyping. Considering and discussing how unidentified cryptic taxa might alter experimental interpretations is essential.
Our literature review shows that cryptic taxa are prevalent, implying that many conclusions related to biodiversity, species traits, and within taxon genetic diversity based at the morphospecies level are likely to be inaccurate. Additionally, there is substantial evidence that such cryptic taxa can be linked by evolutionarily but not ecologically significant levels of gene flow (section 2.5). Failing to recognise cryptic taxa and appropriately adjust interpretations can result in misleading conclusions about fundamentally important aspects of biodiversity measurements in corals, and thus accommodating cryptic taxa will be essential for making informed conservation and restoration decisions into the future (Table 1). Below (3.1-3.6), we discuss the most important manners in which biodiversity can be mischaracterised and its effects on conservation and restoration (see also Chenuil et al. (2019) for similar discussions). We also identify ways in which cryptic taxa and hybridisation could potentially aid restoration (3.5). Figure 5 outlines some possible consequences of cryptic coral taxa to current restoration actions such as direct transplantation and coral gardening (Rinkevich, 1995) and other actions that are being actively researched for managing coral populations to improve their resilience to climate change (National Academies of Sciences, 2019; Anthony et al., 2020; Hein et al., 2020; Bay et al., 2023).
3.1 Underestimates of species diversity & overestimates of population sizes
Biodiversity inventories typically determine counts and abundances of distinct species. Common methods, such as field surveys, are usually based on morphological identification of live organisms (‘morphospecies’) and often struggle to confidently identify nominal species (DeVantier et al., 2020). If evolutionarily distinct taxa such as cryptic genetic groups are not appropriately recognised and delineated within recognised morphospecies using genotype datasets, then total species counts will be greatly underestimated and will bias coral population sizes, species extinction vulnerabilities, and biodiversity valuations. For example, estimates of census population sizes for present-day corals have been recently debated (Dietzel et al., 2021; Muir et al., 2022), as smaller population sizes would imply greater vulnerability to extinction (Dietzel et al., 2021; Muir et al., 2022). Conversely, some rare morphospecies may be hybrids (Richards et al., 2008) and thus the number of rare species might be reduced. Phenotypic plasticity could sometimes upwardly bias species counts, but underestimation due to cryptic taxa is likely more prevalent. Regardless, management assessments are often based on population sizes and abundances, and as such, their predictability and accuracy are reduced when estimates rely on species determined solely by morphological characteristics.
Table 1 - How undetected cryptic species affect biodiversity metrics and management
|
Measure |
Actual value likely to be |
Management implications of undetected, co-occurring cryptic species |
|---|---|---|
|
Species richness |
|
Number of species under management is underestimated Management priorities based on current estimates do not reflect evolutionarily distinct groups |
|
Species range |
|
Geographic distributions could be overestimated Endemic species may not be recognised Underestimated risk of extinction for cryptic endemic species |
|
Species niche breadth & phenotypic diversity |
|
Perceived generalist species may instead be comprised of multiple specialist species Translocated cryptic specialist species may fail to establish in new locations due to phenotype-environment mismatch Species’ tolerances to environmental conditions, including temperature, likely overestimated Extinction risks underestimated |
|
Within-population genetic diversity |
|
In locations where cryptic species co-occur, genetic diversity will be overestimated Adaptation potential may be overestimated |
|
Inbreeding |
|
In locations where cryptic species co-occur, inbreeding will be overestimated Baseline estimates of natural inbreeding will be inaccurate |
|
Population structure (e.g. FST) |
|
In locations where cryptic species co-occur, population structure measures will be inaccurate Gene flow (and dispersal) will either be over- or underestimated depending on proportions of cryptic species |



3.2 Overestimates of species ranges, niche breadths & generalist phenotypes
Trait diversity assessments will be compromised if trait measurement occurs on an amalgamation of distinct taxa. The combined group is likely to contain more variation than the distinct groups, which will inflate estimates of both genetic and phenotypic variance for natural populations. This phenomenon can affect data interpretation for a diverse array of coral traits. For example, species range estimates would be upwardly biased if cryptic species are geographically restricted within the broader range of a presumed ‘species’ (as defined by morphology). Environmental niche breadth can also be overestimated: for instance, many coral species that were previously considered depth generalists could resolve into taxa with more restricted depth distributions when cryptic species are considered (e.g. Bongaerts et al., 2021a). Similarly, a presumed species may appear to have a generalist phenotype but actually comprise multiple taxa that are more specialised, as appears to be the case for bleaching responses among Orbicella faveolata taxa (Gomez-Corrales & Prada, 2020) and thermal tolerance traits among A. hyacinthus taxa (Naugle et al., 2024). This means that cryptic taxa may not be ecologically or functionally equivalent. Thus, restoration actions that involve coral outplanting (i.e., fragments or sexually propagated colonies) risk mismatching the source taxon’s niche with their new destination’s environmental conditions, potentially compromising the growth and survival of the outplants (Edwards et al., 2010; Shaver et al., 2020) (Fig. 5). Erroneous estimates of species niches and phenotypes would similarly undermine the accuracy of species distribution models that might be used in planning to predict future locations with suitable environments under climate change. More studies are urgently needed to test how often sets of cryptic coral taxa differ in their phenotypes, including their preferred niches. Current evidence suggests that conclusions drawn from incorrectly identified taxa are likely to be overly optimistic in terms of species ranges and niche breadth, and therefore will likely overestimate coral resilience to environmental change.
To get a sense of how often unrecognised cryptic coral taxa could affect inferences from other fields in coral biology, we investigated whether thermal biology studies are alert to cryptic coral taxa (Box 3). If cryptic coral taxa consistently differ in their phenotypes, including response to thermal stress, then experimental outcomes need to be evaluated by taxon. Yet, we find that only 8% of such studies included genotyping that could identify cryptic taxa, suggesting that many studies could be inadvertently evaluating multiple cryptic taxa as single species and thereby producing incorrect or biased conclusions. Although it is outside the scope of this paper to examine the thousands of papers in coral biology that examine phenotypes (including niches and geographic distributions), it is likely that most experimental and ecological studies have not genotyped colonies nor applied any criteria to detect cryptic taxa (such as those presented in section 2.1). Thus, inferences describing species attributes from many studies need to be viewed with scepticism, especially if policy and management actions are based on the conclusions of these studies. (We recognise that incorporating genotyping into experimental designs increases costs and efforts, nonetheless, coral biologists should design their investigations with the assumption that cryptic taxa may be present and consider how interpretation may change if their experimental subjects are a mix of taxa – see Box 2 for discussion and suggestions.)
Box 3: Are coral experiments designed to detect cryptic taxa?
Overlooking cryptic taxa can bias interpretations of experimental results. To ascertain how substantial this issue might be for coral studies, we focus on experiments related to thermal tolerance as a subset of coral studies more generally. Marine heatwaves have caused extensive coral mortality events globally (Leggat et al., 2019), and thus numerous coral studies have aimed to ascertain intra- and inter-specific differences in phenotypic and physiological heat stress responses using experiments (e.g. common gardens, reciprocal transplants, etc.) and natural heating events. Mounting evidence suggests that cryptic species display contrasting responses to heat stress (Gomez-Corrales & Prada, 2020; Rose et al., 2021; Rivera et al., 2022; Grupstra et al., 2024), and so experimental results may be more accurate when considering potential cryptic taxa – identified using genomic-scale genotyping.
The Coral Research and Development Accelerator Platform (CORDAP) database (Ortiz et al., 2023) represents a curated search for papers that study thermal biology of corals. We screened the database to identify records which used genome-wide data of the coral host (i.e., coral SNP data) and those that conducted either ordination or model-based clustering (as in 2.1). We evaluated the database to determine:
-
The number of studies within the database undertaking experiments that genotyped corals for multiple unlinked markers (i.e., created data that could be used for ordination or model-based clustering).
-
The proportion of these studies that performed either an ordination or model-based clustering based on individual genotypes.
-
Whether there is evidence for cryptic taxa based on applying the criteria outlined in Section 2.1.
We found that very few experimental studies genotyped coral colonies: from 562 studies, only 60 studies included any sort of host genotyping and only 17 used high-resolution genome-wide markers such as SNPs. Still fewer studies undertook either ordination or model-based clustering on their genomic data (n=8; Fig. T2). For these eight studies, it was essentially impossible to evaluate ordination and model-based clustering outputs for evidence of cryptic taxa (in line with 2.1) because the number of surveyed individuals was so low. Three studies, however, included protocols to detect or pre-select cryptic taxa (Rose et al., 2017; Ruiz-Jones & Palumbi, 2017; Rose et al., 2021). Given that we estimate over 50% of coral studies targeting a single species encounter cryptic taxa (2.1), it is highly likely that hundreds of experimental studies will have inadvertently sampled multiple taxa. Therefore, we would anticipate that reported variances among individuals within studies would be greater than the true variances within cryptic taxa (3.2). This could manifest as an overestimation of thermal tolerance breadth and thus may also mask differences or similarities in measured tolerance in comparative tests between morphospecies. The CORDAP database focuses on one group of studies, but we would anticipate that similar issues arise across all coral experimental work that does not leverage genomic-level genotyping of individual colonies.
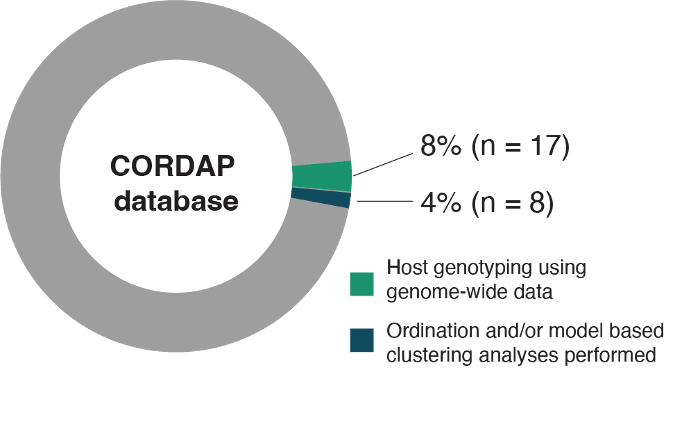
Figure T2 – Proportion of individual studies from the Coral Research and Development Accelerator Platform (CORDAP) thermal tolerance experiment database that: (1) record genotype data capable of identifying cryptic taxa via genome-wide data of hosts, and (2) used ordination-based analyses or model-based clustering analyses.
A major consequence for ignoring cryptic genetic groupings is that common measures of population genetic variation and gene flow are likely to be biased and incorrect when discrete taxa are not accounted for within the analysis (Pante et al., 2015). Lumping genetically distinct groups will inflate apparent within-population diversity based on measures of allelic diversity or expected heterozygosity and overestimate inbreeding via the Wahlund effect (as discussed in Schmidt et al., 2023). When comparing across geographic locations, measures of population structure (notably FST) can be biased either upwards or downwards, depending on the mix of cryptic taxa sampled (Pante et al., 2015). These phenomena are neatly illustrated and discussed by Sheets et al. (2018) for A. hyacinthus in the western Pacific (see also Warner et al., 2015). Similarly, common summary statistics such as genetic diversity, differentiation, inbreeding, and effective population size can be biased when introgression between differentiated taxa is not considered (Hoban et al., 2022). For example, introgression may inflate the measured diversity for populations that include individuals of mixed ancestry, while population structure (FST and other similar measures that use both between and within population variation) can be biased in either direction depending on admixture proportions in populations being compared. In summary, cryptic taxa are likely to greatly affect the accuracy of studies aiming to assess and monitor genetic diversity.
3.4 Diminished reproductive success due to species isolating barriers
The existence of cryptic taxa implies that many coral groups sit somewhere along the speciation continuum with some degree of reproductive isolation between taxa. Intrinsic reproductive incompatibilities would, therefore, be problematic for spawning corals in captivity, as between-taxa crosses could be blocked when potential parents include more than one taxon. In pooled spawning with multiple parents, reproductive blocks between some sets of parents would increase variance in reproductive success among parents and thereby diminish genetic diversity among resultant offspring.
For restoration actions that involve outplanting (regardless of whether outplants are produced sexually or by clonal propagation), intrinsic reproductive incompatibilities between cryptic taxa could be detrimental or beneficial to restoration, depending on the precise goal. For example, a restoration goal may be for outplanted corals (chosen for specific trait attributes such as thermal tolerance) to interbreed with local corals and thus spread advantageous alleles. However, incompatibilities could prevent outplanted colonies from being able to interbreed with colonies in receiving populations or to produce viable and fertile offspring (compounding any effects of phenotype-environment mismatches as described in 3.2). Alternatively, maintaining partial genetic isolation between outplanted and receiving populations of corals could be considered advantageous if the outplanted corals contain co-adapted gene complexes that enhance their fitness or the fitness of F1 hybrids. In this case, diminished effective gene flow – whether genome-wide or associated with inversions that contain co-adapted loci – would protect beneficial combinations of alleles from being separated by recombination (Barton, 2024), thereby allowing the beneficial outplanted gene combinations to persist among the descendants of outplanted corals. Similarly, restoration actions involving manipulated genes – should that technology become feasible for restoration (Cleves, 2022) – would also be affected by reproductive incompatibilities. On the one hand, reproductive isolation between outplants and receiving corals could be viewed as reducing the impact of a restoration action by limiting spread of manipulated genes in the wild, but on the other hand, a natural block to spreading could potentially reduce risks of unconstrained modified gene release in the wild and avoid the spatial dilution of enhanced corals. Thus, it will be important to clarify restoration goals and then consider how interactions between cryptic taxa could modify intended outcomes.
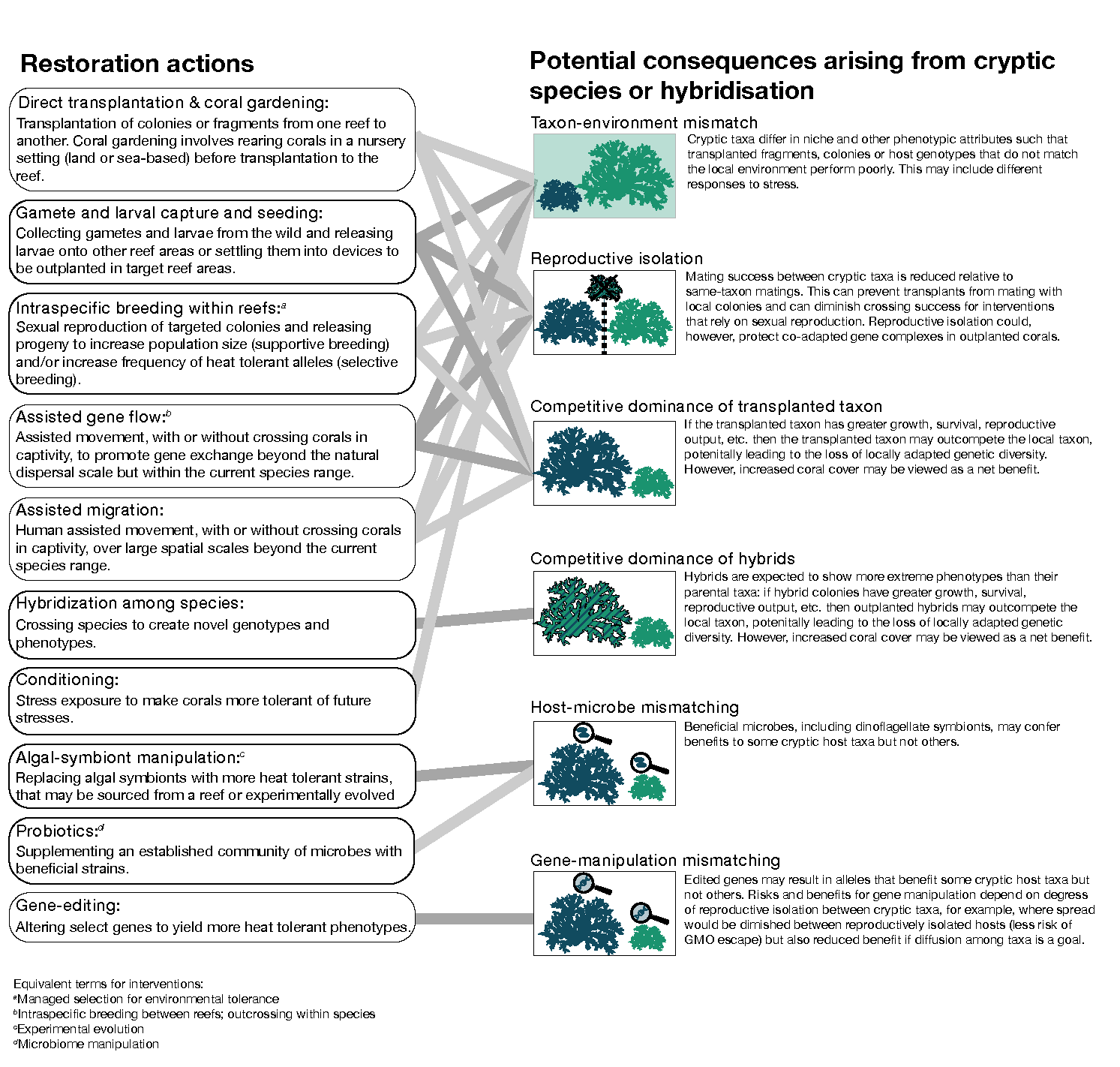
Figure 5 - How cryptic coral taxa and hybridisation could affect coral reef restoration actions that aim to preserve biological diversity, counter population declines, and/or promote resilience to climate change through biological adaptation. Terminologies follow (van Oppen et al., 2015; National Academies of Sciences, 2019; Hein et al., 2020; Bay et al., 2023) and are not mutually exclusive. For example, assisted movement could be undertaken on fragments (direct transplantation), larvae, or via the progeny of captive, sexually propagated corals. In all graphics, the coral colony shown as a larger size indicates greater fitness (e.g. survival, reproductive output, thermal tolerance) leading to competitive dominance. Different shades of grey lines are used to aid visualising connections but do not convey any specific meaning. Within images, blue coral silhouettes indicate translocated colonies whereas green silhouettes indicate local colonies.
3.5 Evolutionary consequences arising from hybridisation
Because first generation (F1) hybrids often have higher relative fitness (Mackay et al., 2021), cross-breeding species has been proposed as a possible coral restoration strategy (National Academies of Sciences, 2019; Bay et al., 2023). Towards this end, nominal species of Acropora have been crossed and the performance of hybrids evaluated against parental species under laboratory (Chan et al., 2018; Chan et al., 2019) and field conditions (Willis et al., 2006; Fogarty, 2012; Lamb et al., 2024). Encouragingly, none of the experiments indicated lower fitness of hybrids, however, hybrids were also not universally superior to parentals across various fitness-related traits. Experiments based on fragmenting adult colonies morphologically identified as parental and F1 also found no survival difference between groups but did find that putative F1 fragments grew faster (VanWynen et al., 2021). If first-generation hybrids have higher fitness than parental species under field conditions, then their enhanced performance could be viewed as a restoration boon (better surviving coral colonies) or a liability (hybrids outcompete parental species) depending on restoration goals (mirroring considerations under 3.4).
However, it is well-known that backcrosses and all of the possible variations of later generation hybrids can yield much greater phenotypic variation than their parental species (i.e., transgressive segregation, Abbott et al., 2013; Mackay et al., 2021) and recent simulations indicate that such hybrid populations can rapidly adapt to novel environments not previously experienced by either parental species (Kulmuni et al., 2024). Thus, coral populations that have experienced past hybridisation may be best suited for future rapid adaptation. Additionally, there may be opportunities to develop restoration strategies using naturally occurring hybrids. For example, locating early and late-generation hybrid colonies in wild populations and characterizing their spatial and environmental distributions could potentially help source coral colonies with heat-resistant or other adaptive phenotypes.
3.6 Summary: conservation and restoration research and planning cannot afford to ignore cryptic taxa and hybridisation
Overlooking cryptic taxa can yield inaccurate conclusions about species’ abundances, ranges, niches, phenotypic variance, and patterns of within-species gene flow and genetic diversity (Table 1). Conservation and restoration plans based on such erroneous estimates of species attributes are unlikely to achieve their goals (3.1, 3.2). Therefore, we urge coral biologists to acknowledge cryptic taxa as an important source of bias (Box 2) and ideally seek to minimise this error source in their investigations (Box 3). Development and implementation of coral restoration methods will similarly need to characterise cryptic taxa, where loss of genetic diversity under crossing experiments is especially concerning (3.4).
And yet, the prevalence of cryptic taxa suggests that coral biodiversity is much richer than anticipated and therefore some taxa may be inherently resilient to future conditions and/or contain sufficient genetic variation for rapid adaptation. Gene flow between taxa appears to be a common attribute of coral biology and this characteristic may provide mechanisms for adaptive traits to spread between taxa and to develop new restoration strategies capitalising upon pre-existing advanced generation hybrids (3.5).
Corals present a largely overlooked system for studying speciation and adaptation, despite extensive evidence for recent cryptic divergence. Studies of coral speciation and diversification processes can advance understanding of how coral biodiversity emerges and is maintained. Such studies would, in turn, characterise biological attributes that are important for reef conservation management decisions and strategies. Here we highlight several topics worthy of focused study.
The emerging consensus that closely related coral taxa are frequently sympatric at coarse spatial scales yet segregate by depth or other microenvironmental characteristics aligns well with models of ecological speciation (Schluter, 2001; Rundle & Nosil, 2005). Furthermore, the presence of distinct cryptic taxa in close geographic proximity suggests that selection for microhabitat matching (such as depth) may be very strong (a high selection to migration ratio: Richardson et al., 2014) and/or that intrinsic reproductive barriers enhance genetic differentiation between habitats (Bierne et al., 2011). To what extent cryptic taxa differ phenotypically or in terms of competitive ability is unknown, although differences in bleaching susceptibility among some cryptic taxa suggest differing vulnerabilities to climate change (Gomez-Corrales & Prada, 2020; Rose et al., 2021; Rivera et al., 2022).
It is likely that intrinsic barriers to reproduction limit gene flow between taxa to some extent. Analyses to date support evolutionary genomic models that allow genomic regions to differ in permeability to gene flow (see section 2.5 and Fig. 4), which may be consistent with chromosomal inversions or other genome features contributing to reproductive isolation. More studies that use chromosomal resolution genotyping will be critical to forming a deeper understanding of how species boundaries are maintained in corals (e.g. Leitwein et al., 2020) and can guide decisions on assisted migration or choosing broodstock for selective breeding (Fig. 5).
Although the relative importance of extrinsic (including environmental) and intrinsic barriers to reproduction are undetermined, as sessile organisms, corals are well-suited to manipulative experiments. Experimental designs based on common gardens and reciprocal translocations can provide some of the strongest tests of local adaptation and therefore provide evidence for environmental selection (Kawecki & Ebert, 2004). Additionally, the clonal nature of corals means that genetically identical fragments from the same colony can be exposed to differing treatments, offering rich opportunities to combine experiments with genomic analyses to holistically investigate the interactions between taxon identity, phenotype, and environment (Pinsky et al., 2023; Richards et al., 2023).
Divergence dates between cryptic taxa often pre-date Holocene reef configurations (Fig. 4), implying that old standing genetic diversity is spread across contemporary reefs that are characterised by spatially complex yet replicated microhabitats and environmental gradients. Thus, corals are ideal for investigations that explore the genetic mechanisms of parallel divergence, especially over depth gradients (e.g. analogous to fishes that have spread into post-glacial lakes: Rougeux et al., 2017; De-Kayne et al., 2022). Such investigations would also provide insights on the geographic distribution of standing genetic variation, which may be under increasing selective pressure due to pervasive anthropogenic environmental changes. For example, knowing whether geographically distant populations do or do not share alleles for advantageous traits can guide decisions regarding the utility of assisted gene flow. This is because evolutionary rescue is only worth considering if donor and recipient populations have different standing genetic diversity for ecologically functional traits.
Individual colonies with genotypes consistent with recent hybrid ancestry have been noted in the current literature (section 2.5). To date, these likely admixed individuals have primarily been documented as anecdotal observations rather than being the focus of detailed studies. However, research on hybridisation and hybrid zones offers valuable insights into speciation and adaptation, highlighting the need for more focused investigation of these individuals (Hewitt, 1988; Harrison, 1990). Potential restoration interventions based on hybridisation rest on the supposition that hybrid corals differ in their phenotypes relative to parental species – due to some combination of hybrid vigour or transgressive segregation (section 3.5). Yet, aside from the transect studies of Prada & Hellberg (2014), no other study has mapped the spatial and environmental distributions of hybrids relative to parental taxa nor compared their phenotypes. For example, finding and characterising the phenotypes of advanced generation hybrids (not just F1’s) would enable robust tests of hybrid fitness and evaluate evidence for transgressive segregation. Integrated field and genetic studies could thus be critical for advancing our understandings of coral hybrid zone dynamics (see Westram et al., 2018 for a marine example).
Throughout this review, we have focused primarily on the cnidarian component of coral genomes to document evidence for cryptic species and hybridisation. However, in considering how future studies could build on these observations to better understand speciation and adaptation processes, it will also be important to integrate genetic analyses of the coral host as well as the associated symbiotic dinoflagellates and microbial communities. An exciting line of investigation would be to try to understand the co-evolutionary dynamics of hosts and symbionts in reference to environmental adaptation and speciation, where environmental heterogeneity likely exerts direct selection on the genomes of both corals and their symbionts (i.e. the coral holobiont) and indirect selection via host-symbiont genetic interactions.
In this review, we demonstrate that cryptic coral taxa are extremely common and are often connected by low levels of gene flow. Although our assessments reflect findings for shallow-water corals, we would anticipate that deep-water corals also harbour substantial cryptic diversity. Our failure to locate population genomic studies of deep-water corals indicates that more genetic studies of deep-water corals are needed. The prevalence of cryptic coral taxa among shallow-water corals means that many accepted understandings and conclusions regarding coral biology could be incorrect. In Section 3, we highlight how ignoring cryptic taxa can mislead management decisions by: biasing estimates of spatial biodiversity patterns; inflating species home ranges, trait spaces, and niches; and skewing inferences regarding intraspecific population structure and gene flow. Thus, as a field, we are unable to confidently generalise species distributions, ranges, and phenotypes including resilience to heat stress without genotype-based analyses to adequately assess the potential for cryptic coral taxa (Box 3). Identifying locations with high genetic diversity that may harbour greater adaptive potential or inferring locations with high gene flow and dispersal will crucially depend on analyses that are able to detect and account for distinct taxa. Without genomic and/or experimental data on hand, a precautionary principle may be to assume that populations inhabiting distinct environments (especially depth) are likely to be evolutionarily distinct and ecologically independent.
Although observations of cryptic coral taxa are frequent, our collective knowledge regarding the evolutionary dynamics that enable closely related taxa with incomplete species boundaries to persist in sympatry remains limited. There is a vast potential to unite coral studies with the insights and approaches from studying speciation and adaptation from other fields and organisms (section 4). For example, partnerships between coral ecologists, physiologists, and population geneticists may bridge insights into microevolutionary responses to climate change, while collaborations with experts from other fields may broker novel analyses and genomic-based approaches to better understandings of speciation and hybridisation in corals.
The reality, however, is that the future for corals and coral reefs is perilous (Knowlton et al., 2021). Management decisions cannot wait for perfect information. An important next step will be for evolutionary biologists to investigate how conservation and management actions can best proceed with a renewed expectation that coral species boundaries are unlikely to be well defined – a conservation challenge that ultimately afflicts many other taxa in addition to corals (Hey et al., 2003; Roux et al., 2016).
Appendices
Structured review for population genomic studies
An initial search of Web of Science Core Collection was performed on 21/10/2022, using the search terms “(TI=(coral) OR TI=(scleractinia*) NOT TI=(fish)) AND (AB=(rad*) OR ALL=(snp*))”. The search returned 803 studies. Titles and abstracts were filtered to exclude irrelevant studies. Studies appearing to contain population genomic data of scleractinians or octocorallians were retained for manual inspection. On 4/11/2022 the above search was repeated allowing for “coral” to be in the abstract rather than title and this yielded 853 additional studies for a total of 1620 unique papers that were reduced to 99 after skimming titles and abstracts.
Each manuscript was read and evaluated by two people independently to ensure that genomic data: i) pertained to the cnidarian coral host, ii) used many loci on a genomic scale (i.e., not microsatellites, not metabarcoding), iii) surveyed two or more sites or habitats, and iv) presented results that included ordination analysis based on individual genotypes (i.e., principal components analysis (PCA), principal coordinates analysis (PCOA) or multidimensional scaling (MDS)) and/or unsupervised model based clustering tests (such as ADMIXTURE, STRUCTURE, fastSTRUCTURE or sNMF). We did not consider papers only reporting discriminant analysis of principal components, as DAPC finds the eigenvectors that best differentiate prespecified groups. In contrast, PCA, PCOA and MDS find eigenvalues that best capture total diversity regardless of group membership (see Thia, 2022 for further discussion). If these four conditions were not met, the study was excluded. For each retained study, two evaluators independently extracted key attributes and reconciled discrepancies between their scoring through discussion. Despite attempting to undertake a rigorous and inclusive search, we noticed that several suitable manuscripts were missing and therefore on July 18, 2023 we ran an ad hoc search in Web of Science based on authors that are known to be publishing on population genomics of corals (namely Barshis, DJ; Baums, IB; Bay, LK; Bongaerts, P; Cooke, I; Matz, MV; Palumbi, SR; Richards, ZT, Underwood, JN, van Oppen, MJH) and also repeated the above search with exactly the same criteria for articles published since 21/10/2022. These new papers were evaluated as above. The ad hoc search initially identified 897 papers that were reduced to 16 papers once titles and abstracts were skimmed. In total, 41 papers were found suitable for data extraction representing a total of 51 records (unique paper by species combinations).
Heat stress studies
The CORDAP database (Ortiz et al., 2023) was downloaded on 11/09/2023 and searched for any records which used genome-wide data of the coral host (i.e., coral SNP data) and those which conducted clustering analyses (as described above). First, records were filtered based on whether the database columns “Host_genotype”, “Symbiodiniaceae_genotype”, and “Microbiome_genotype” were listed as “TRUE”. Second, the columns “Symbiodiniaceae_genotyping_approach”, “Host_genotyping_approach” and “Microbiome_genotyping_approach” were interrogated and only records where the genotyping method was listed as reduced representation sequencing (e.g. Restriction site Associated DNA sequencing), whole genome sequencing (WGS) or RNA sequencing (RNAseq) were kept for further checks. The titles and abstracts of the remaining records were checked, and only records for which host genotyping was performed were included. The number of cryptic species assigned in each paper by the original authors was noted, as well as the evaluators’ interpretation of the number of cryptic species based on the plots and analyses (following guidelines in 2.1).
The original database consisted of 562 records, many of which did not include host, Symbiodiniaceae, or microbiome genotyping. The initial filtering of the database for records that included some aspect of host genotyping yielded 222 results and was reduced to 17 records that included genome-scale genotyping. Of these 17 studies, nine studies included either ordination or model-based clustering analyses (e.g. PCA, ADMIXTURE). Of these, three (Rose et al., 2017; Ruiz-Jones & Palumbi, 2017; Rose et al., 2021) matched the criteria in Section 2.1 showing evidence for cryptic taxa. The remaining six studies were either ambiguous or showed no clear evidence for cryptic taxa.
Acknowledgements
We thank M.J.H. van Oppen, K. Johannesson, R. Pears, L. DeVantier, E. Pante, and two anonymous reviewers for their comments and valuable suggestions. S. Starko generously shared photographs of Porites sp. Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Evolutionary Biology (https://doi.org/10.24072/pci.evolbiol.100783; Pante, 2024).
Funding
This work was supported by the Reef Restoration and Adaptation Program, funded by the partnership between the Australian Government’s Reef Trust and the Great Barrier Reef Foundation. CR was supported by a guest professorship (“Marks professur”) funded by Göteborgs Universitet during part of the manuscript preparation.
Conflict of interest disclosure
The authors declare that they have no financial conflicts of interest in relation to the content of the article.
Data, scripts, code, and supplementary information availability
Data, scripts and code are available online (https://doi.org/10.48610/76456b1; Riginos et al., 2024).