
 CC-BY 4.0
CC-BY 4.0
Context and general description
In addition to being a shared natural heritage that we have the responsibility to protect, marine seagrasses provide us with a wide range of essential functions. Seagrass habitats have been described as one of the most productive temperate coastal habitats and are a key component of coastal areas (e.g. Unsworth et al. 2022). As ecosystem engineers, they provide a nursery habitat for many commercial species, filter estuarine water, protect from coastal storms, and sequester pollutants (Larkum et al. 2006). These habitats are however among the most threatened marine ecosystems on Earth, having declined by 29% globally since the 1980s (Orth et al. 2006; Waycott et al. 2009). Despite the implementation of the European Habitats Directive in 1992, most listed EU marine habitats, including seagrasses, were still in a poor or bad conservation status in 2020 (EEA 2020). The European Life project MARHA (MARine HAbitats) aims to improve knowledge of marine habitats, their ecological functions and the pressures they face. Within this framework, the "Functionality of seagrass meadows" work package aims to evaluate the ecological functions provided by Zostera marina Linnaeus, 1753 and Zostera noltei Hornemann, 1832, two emblematic habitat-forming species, throughout their distribution range along the French coast. The dataset presented includes all observations made at four sites spread throughout the range of the species in France over five seasons. The local heterogeneity of these habitats was also explored by sampling at each site in the historically known area of the meadow, in the more dynamic area where the meadow presence has fluctuated in the past, and in the adjacent bare area (hereafter modalities). In addition to a detailed description of Zostera spp. meadows in these three modalities, two main functions of seagrass meadows were studied: the specific diversity of the associated benthic communities (epifauna and suprabenthos) and the benthic fluxes (organic carbon storage capacity, oxygen and nutrient recycling). This comprehensive dataset provides baseline data that can inform science, management and policy. The author has chosen to provide a graphical representation of each dataset and a brief description in order to facilitate visualization and appropriation of the data by readers and potential data users.
Experimental design and sampling procedure
Study area description and study extent
We carried out field surveys of Zostera spp. in four selected sites along the English Channel, Atlantic and Mediterranean coasts (Fig. 1). The four sites are part of the “Natura 2000” sites of the European “Habitat Directive” network and host monospecific Z. marina and Z. noltei beds.
Dinard (DN) is the northernmost site. Located in northern Brittany, this megatidal site is open to the English Channel. The Z. noltei beds are intertidal, and cover about 22 ha, whereas the Z. marina beds are mostly subtidal, with some possible short periods of emergence during spring tides. Further south, the Golfe du Morbihan (GM) is a 11 500 ha semi-enclosed sea (mesotidal) open to the Bay of Biscay through a < 1km channel with powerful tidal currents (up to 22 m/s during ebb tides). The sampled seagrass beds were located in the eastern part of the gulf in a relatively sheltered area. The Z. noltei beds are intertidal and the Z. marina beds are subtidal. Arcachon Bay (AC) is a 17 400 ha mesotidal enclosed bay on the Atlantic coast. It is connected to the Atlantic Ocean by a channel that is 2 to 3 km wide. Its intertidal area (11 700 ha) is covered by mudflats, partly occupied by Z. noltei and oyster beds. Its subtidal area (5 700 ha) is partly colonised by Z. marina. The Thau lagoon (TH), located on the Mediterranean coast, has an area of 7 500 ha and is relatively deep (average depth 4.5 m). The Zostera beds are mainly located in the southern part of the lagoon. The site is microtidal and the seagrass beds are always submerged.
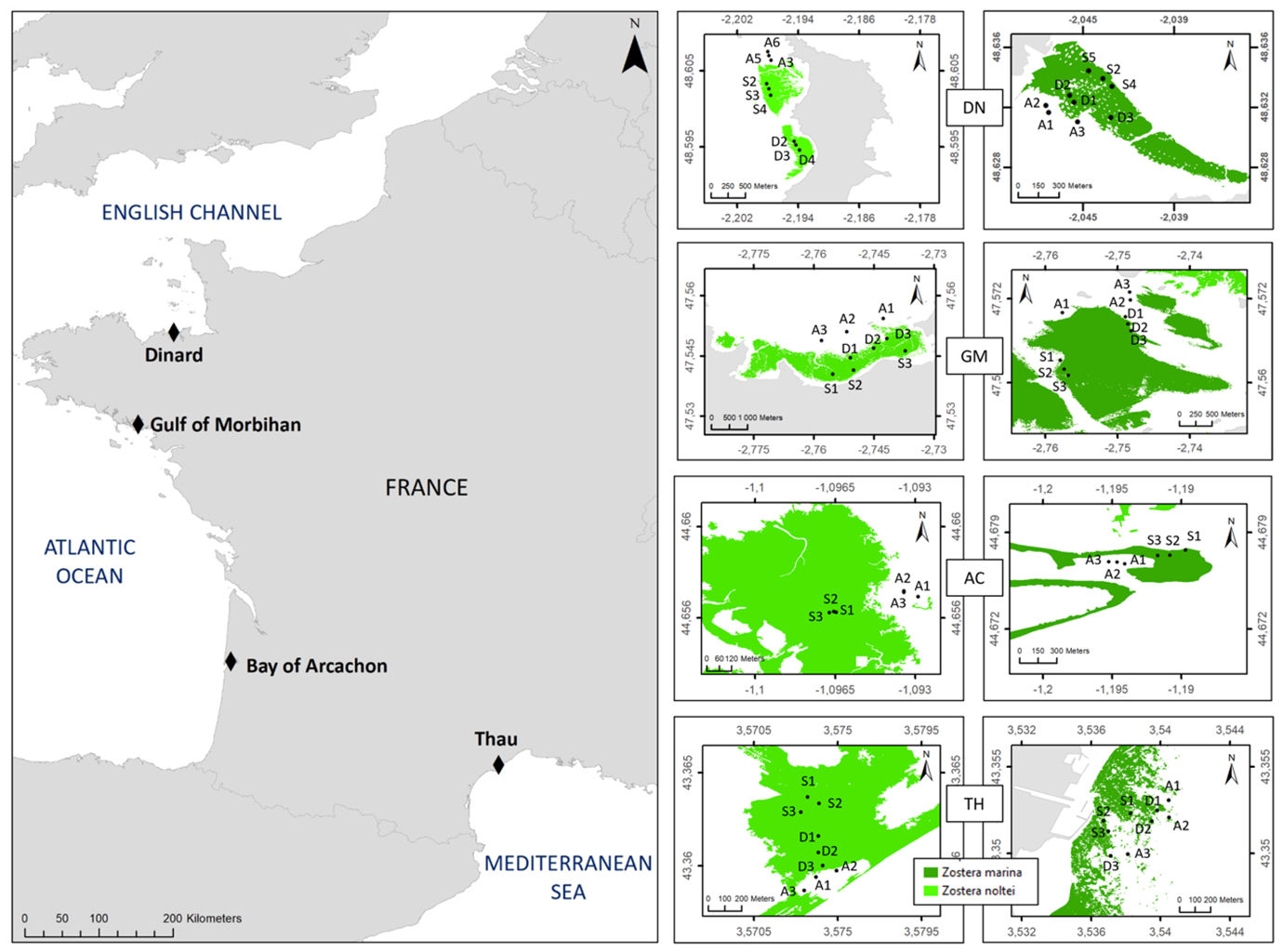
Figure. 1 - Location of the four study sites along the French coasts with positioning of the sampling points in Zostera marina (dark green) and Zostera noltei (light green) habitats. DN: Dinard, GM: Golfe du Morbihan, AC: Arcachon and TH: Thau ; Modalities: A: bare, S: stable, D: dynamic.
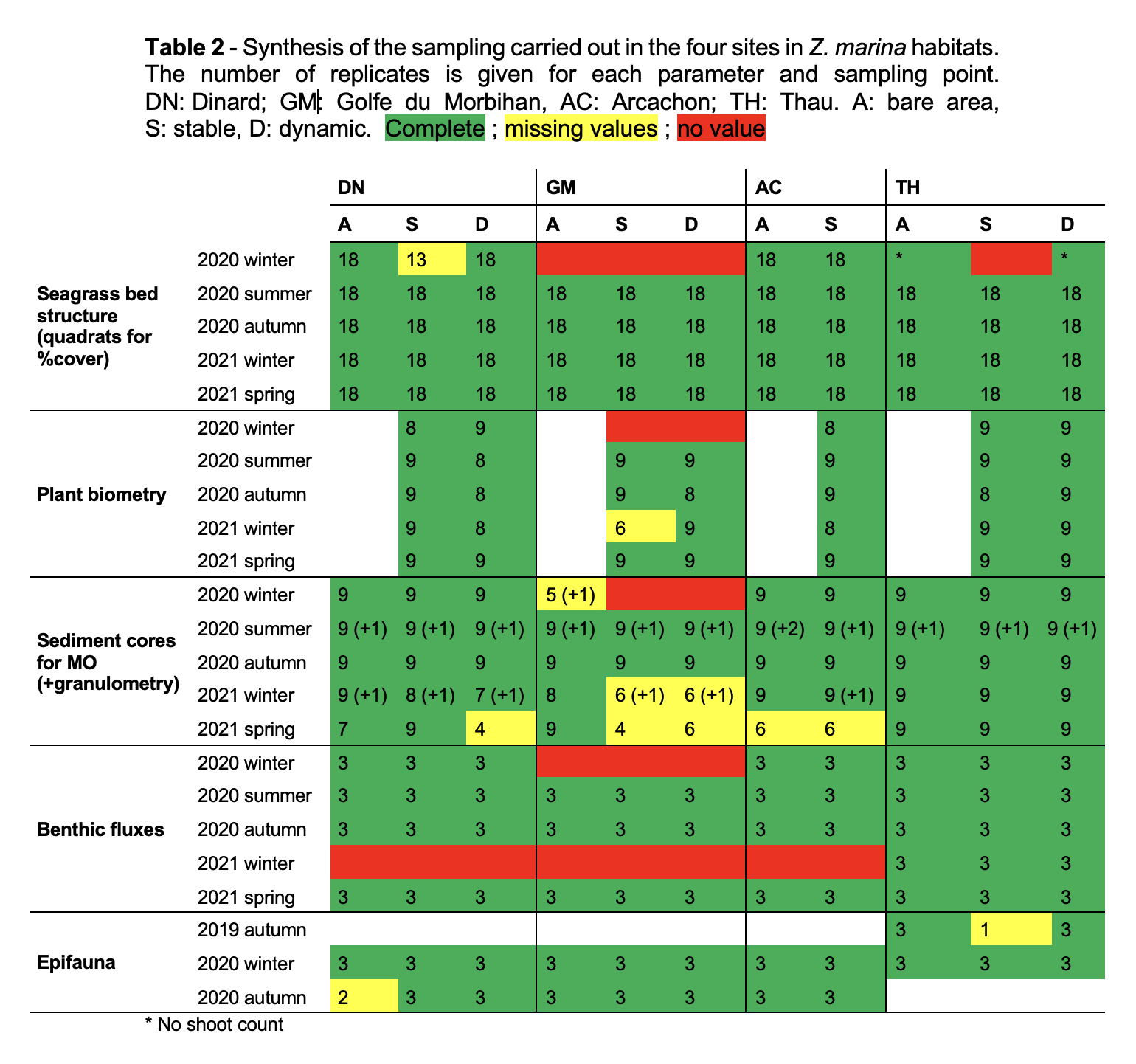
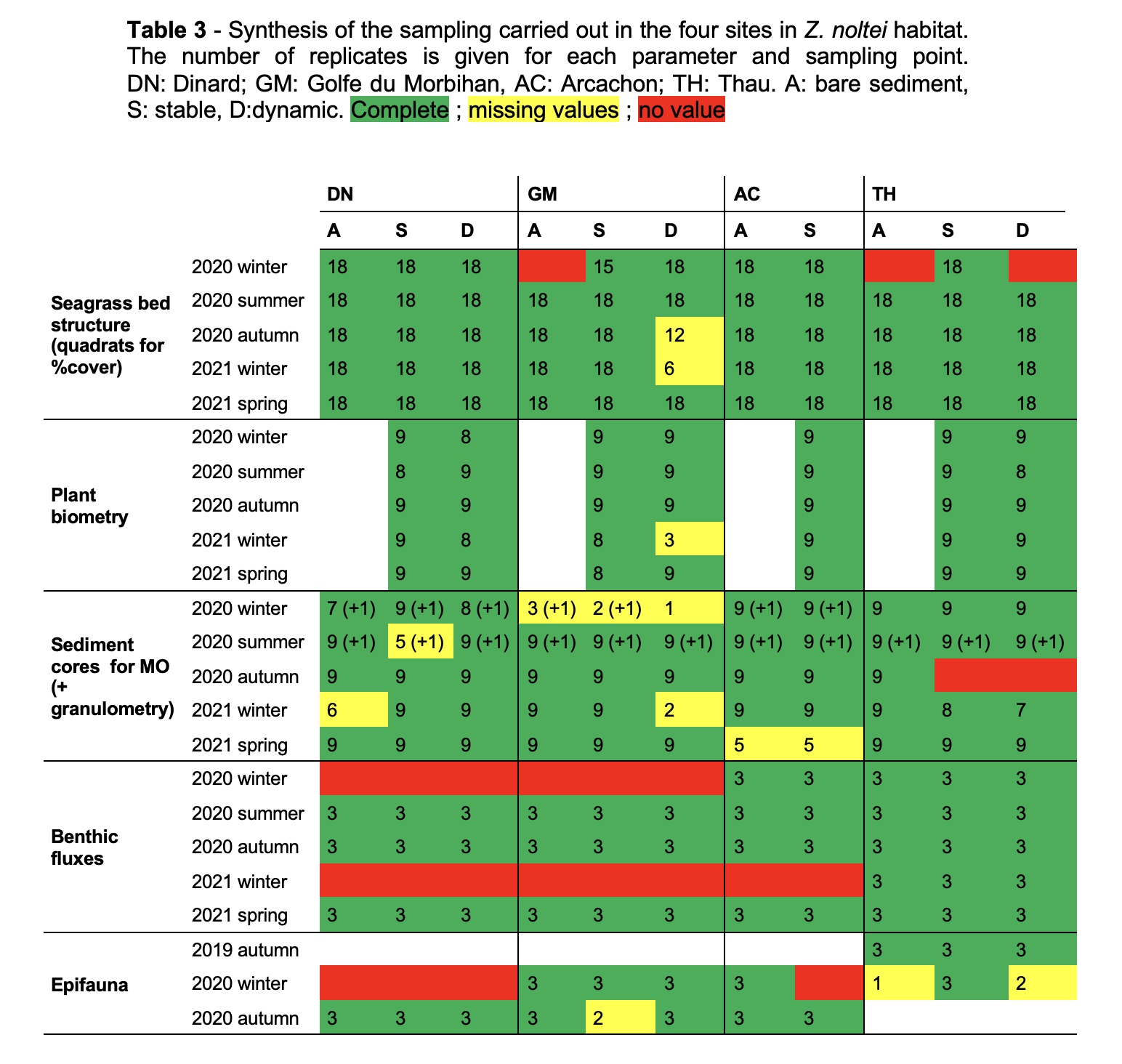
Three spatio-temporal modalities were defined for each Zostera habitat (Fig. 1), as gathered from historical maps and/or area photographs where available, or on local expert knowledge: a bare modality close to the seagrass bed where no evidence of Zostera spp. was found over the last decade (A), a stable modality where the seagrass bed has been observed continuously over time (S), and a dynamic modality where the seagrass bed has been observed recently but where records showed signs of absence (D). This last modality corresponds to the most recently colonized habitat. Three stations were placed in each of these three modalities. From three to six samples were taken at each station, depending on the parameter of interest (see next paragraphs). In AC, no distinction could be made between the two modalities due to the specific configuration of the seagrass beds which have been in sharp decline since 2000 (Plus et al., 2010).
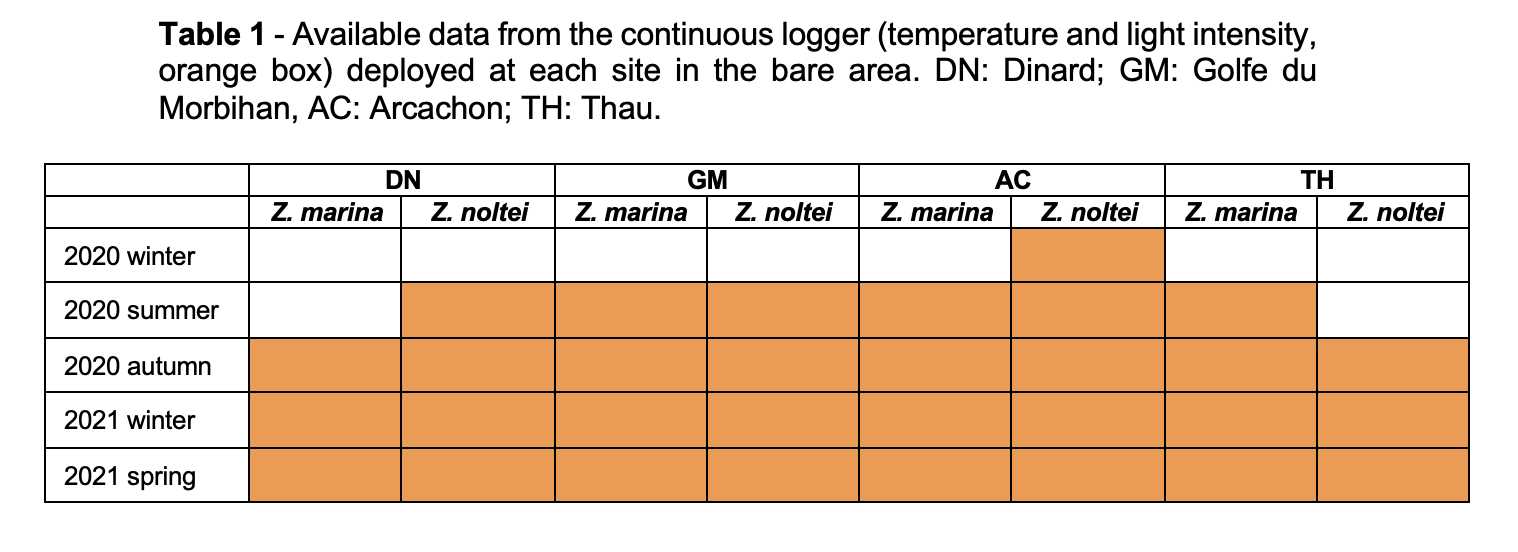
The project included between two and five sampling campaigns (Tables 1, 2 and 3), conducted from autumn 2019 to spring 2021, with some hiatuses in spring 2020 due to the COVID-19 pandemic, and in winter 2020 and 2021 due to meteorological conditions. See Tables 1, 2 and 3 for the full inventory of available data.
Environmental data
High frequency (10 min sampling rate) loggers (HOBO® Pendant® UA-002-64 and (HOBO®-U20 L, accuracy 1 cm, resolution 0.21 cm) were placed on each bare sediment area (modality A) approximately 0.1 m above the seabed, to measure light intensity (lumens m-2), temperature (°C) and tidal water level fluctuations (cm) (except in TH).
Seawater was sampled once during each seasonal campaign, near the stable modality, to estimate salinity, chlorophyll-a (chl-a) and nutrient concentrations. Nutrient concentrations were quantified by segmented continuous flow analysis using colourimetric (NOX and PO43-) and fluorimetric (NH4+) detectors (Aminot and Kérouel 2004). Chl-a was estimated from 90% acetone extraction of filtered seawater, by spectrofluorometry (Neveux and Lantoine 1993).
Three sediment samples were collected once during each seasonal campaign in each station (N=3 per station, N=9 per modality) for organic matter analysis. The percentage of organic matter was determined by the loss on ignition method after 6 hours at 520°C.
Additional sediment samples were taken in winter and summer for each modality (N=1) where possible for grain size analysis. Sediments were oven dried (24 h at 60°C) and separated into 13 fractions (<40 μm, 40, 50, 63, 100, 125, 150, 250, 400, 500, 630, 800, 1000, 2000) whose masses were measured. The percentage of each fraction was calculated from the total weight of the sample.
Seagrass bed structure and plant biometry
Leaf cover in vegetated areas was estimated visually by divers or by fieldworkers at low tide, from six quadrats (0.16 m2) at each station (N=18 per modality). For Z. noltei, in TH (submerged plants), a PVC sheet was used to flatten the leaves on the sediment. For Z. marina, all shoots in the quadrats (or a sub-sample in the case of high cover) were counted to express the density per m2.

Figure. 2 - Suprabenthic sledge in use during a field campaign. © IFREMER
Three PVC cores (0.005 m2 for Z. noltei, 0.03 m2 for Z. marina) were collected in each station. Each core was sieved (1 mm), and macroalgae and shoots were carefully washed with filtered seawater to remove epiphytes and sediment. Shoots were counted and fully mature leaves were counted and measured manually (length and width of each leaf) for five randomly selected shoots per replicate. The total number of shoots per core was used to estimate the density of Z. noltei. The leaf area (length × width of all measured leaves) was calculated per core and expressed in m2. Finally, the dry mass of leaves, roots and rhizomes of seagrass and macroalgae was estimated for each core after freeze-drying.
Epifauna
In winter and autumn 2020 (except for Thau: autumn 2019), epifauna and fish were sampled using a suprabenthic sledge of 1 m width equipped with a 1 mm mesh net (Fig. 2). Sampling was carried out over a 25 m transect, corresponding to a total sampled area of 25 m2. Three transects were conducted in each site and modality (A, S, D), resulting in a total of N=9 transects per site to cover habitat heterogeneity. Epifauna and fish were preserved in ethanol (96°C) until identification to the lowest taxonomic level (species for most of the time). Biomass of each taxonomic group was estimated after combustion at 550°C for 4 h and expressed as AFDM (ash free dry mass in mg). Abundance and biomass were standardized to 1 m2 area.
Benthic fluxes and associated macrofauna
Underwater benthic metabolism was estimated by measuring oxygen, carbon and nutrient fluxes during successive light (under maximum light conditions, both primary production and respiration processes) and dark (respiration) in situ incubations. The benthic chamber method was used, as is common for benthic fluxes measurements (e.g. Smith et al. 2009, Ouisse et al. 2014, Grenz et al. 2021). Three benthic chambers (volume=32 L, surface=0.13 m2, Fig. 3) were simultaneously deployed in each modality (N=3) during each campaign. The heterogeneity of the habitat was deliberately taken into account in our sampling by randomly placing the benthic chambers in each area. The seagrass habitat was therefore considered in all its complexity, with its associated flora and fauna. The incubation time varied from 1.5 to 2.5 hours depending on the site and the season. A submersible pump connected to waterproof batteries ensured continuous gentle mixing of the water in the chambers during incubation. Salinity and underwater photosynthetic photon flux density (PPFD, μmol m-2 s-1) were measured continuously (2 min frequency) approximately 30 cm above the sediment surface using a probe (AQUALABO-C4E) and a spherical underwater quantum sensor (LI-COR193), respectively. Oxygen and temperature were continuously recorded inside chambers (HOBO ® U26-001, 1 min frequency). Water was sampled at the beginning and the end of the incubation for pH and alkalinity measurements. Additional water samples were taken at the beginning and the end of the light incubation for nutrient analysis. pH was measured on board using a multi-parameter probe (WTW® Multi 3620 IDS) equipped with a pH sensor (WTW® IDS SensoLyt 900-P). Alkalinity (mol.kg-1) was measured by pH-metric titration using the Titroline 7000 (SI Analytics, Germany), according to Dickson et al. (2007). Nutrient concentrations were quantified as previously described (§ Environmental data).
The dissolved inorganic carbon (DIC) concentration at each time point was calculated from the pH, total alkalinity (TA), temperature, pressure and salinity using the Seacarb package in R (Gattuso et al. 2023). Finally, flux were estimated from the slope of linear regression for oxygen or from the difference between the initial and final concentrations for nutrients and carbon, expressed per unit of time and surface area. Community respiration (CR, dark incubations) and Net Community Production (NCP, light incubations) were used to calculate the Gross Community Production as follows: GCP = |NCP| + |CR|. The data have been pre-processed according to the rules and corrections listed in Table 4.
Table 4. Corrections applied to the O2 and DIC flux data according to fixed rules.
|
Measured fluxes |
Rules* |
Correction applied if rule broken |
Nb cases |
|---|---|---|---|
|
O2 |
CR <=0 (no production of oxygen at obscurity) |
CR=0 |
<2% |
|
NCP >= CR |
NCP=-CR (→GCP=0) |
3.5% |
|
|
DIC |
CR >= 0 (no consumption of carbon at obscurity) |
CR=0 |
21% |
|
NCP <= CR |
NCP=CR (→GCP=0) |
11% |
* In case of two broken rules, GCP was not calculated (DIC: 6.6% ; O2:<2%).

Figure. 3 - Benthic chamber in use during light incubation. The same model with a dark dome was used to measure respiration. ©IFREMER
Database structure
The dataset is composed of three description tables and eight data tables:
The “Table Description” gives sample codes and metadata on the variables used in the following tables.
The “Sampling Period” table shows the start and end dates of sampling periods for each site and season.
The “Sampling location” table gives the geographical coordinates of each sampling station.
The “environment” table provides information on the environmental parameters at each site and season for the two species (Z. marina and Z. noltei). The tidal fluctuation file is available on request, but is not included in the database due to its size.The “sediment” table indicates sediment organic matter content at the station scale for each season, and granulometry (% of each fraction) in winter and autumn.
The “seagrass_quadrat” table indicates the in situ measurement of Zostera sp. cover (%) and shoot count for Z. marina.
The “seagrass_core” table gives information on the morphometric parameters measured in the laboratory.
The “epifauna_density” table presents values expressed in individuals per m2.
The “epifauna_biomass” table compiles values of ash free dry mass in mg per m2.
The “benthic_fluxes” table gives values of nutrient, carbon and oxygen fluxes with associated environmental conditions : salinity, temperature and PPFD.
The “benthic_fluxes_macrofauna” table indicates values expressed in individuals per core.
Quick data description
Environmental data
Fig. 4 summarizes some of the results from the “environment” table. Dissolved inorganic nitrogen (DIN) concentrations varied between 20 and 60 µmol.L-1, with peaks > 100 µmol.L-1 at some sites during winter. The exception was TH, where concentrations (DIN) never exceeded 1 µmol.l-1. DIN concentrations were mostly due to the pool nitrates + nitrites (NOX). The mean salinity was 31 PSU for the sites of AC, GM and DN and always > 35 PSU in TH. Mean water temperature was close to 9°C in winter and 20°C in summer at all sites, with the highest amplitude in TH (< 8°C in winter and > 25°C in summer). Light intensity in the water varied from less than 200 lumens m-2 to more than 10 000 lumens m-2 for periods of higher intensity.
Seagrass bed structure and plant biometry
This section and Fig. 5 and 6 summarize data from the “seagrass_quadrat” and “seagrass_core” tables. Fig. 5 illustrates the heterogeneity of the seagrass habitat. Sampling revealed areas largely dominated by seagrasses, although macroalgae were also an important part of the habitat, as in GM and TH.
The shoot density of Z. marina could exceed 750 shoots.m-2 but was mostly below 100 shoots.m-2, with seasonal variations. Densities of Z. noltei were one order higher, with values up to > 6000 shoots m-2. Shoot biomass varied between 100 and 400 g m-2 most of the time for both species with site and seasonal variations.
The number of leaves per shoot varied between two and six for Z. marina, and were mostly around two for Z. noltei. For both species, leaf size decreased in winter.
Epifauna
A total of 329 taxa were collected during the study: 119 were common to both Z. marina and Z. noltei habitats, 156 were only associated to Z. marina and 54 solely associated to Z. noltei habitat.
The predominant species in the Z. marina habitat were the gastropods Rissoa membranacea (J. Adams, 1800) and Bittium reticulatum (da Costa, 1778). The maximum species richness observed in one sample was 107 taxa and the maximum abundances were 22 and 56 individuals.m-2, for R. membranacea and Rissoa parva respectively.
In the Z. noltei habitat, the most abundant taxa were R. membranacea and Gobiidae. The highest species richness recorded was 52 taxa, found in only one sample, and maximum abundances were for the gastropod Peringia ulvae (Pennant, 1777) and the amphipod Gammarus insensibilis Stock, 1966 (32 and 15 individuals.m-2 respectively).
Biomass and density were highly variable depending on the site, the season and also the position within the meadow (i.e. Modality, Fig. 7). Maximum mean values were about 700 ind m-2 for density and about 2000g ash free dry mass per m2 for biomass. Lowest biomass and density were near 0. These data can be found in the tables “epifauna_density” and “epifauna_biomass”.
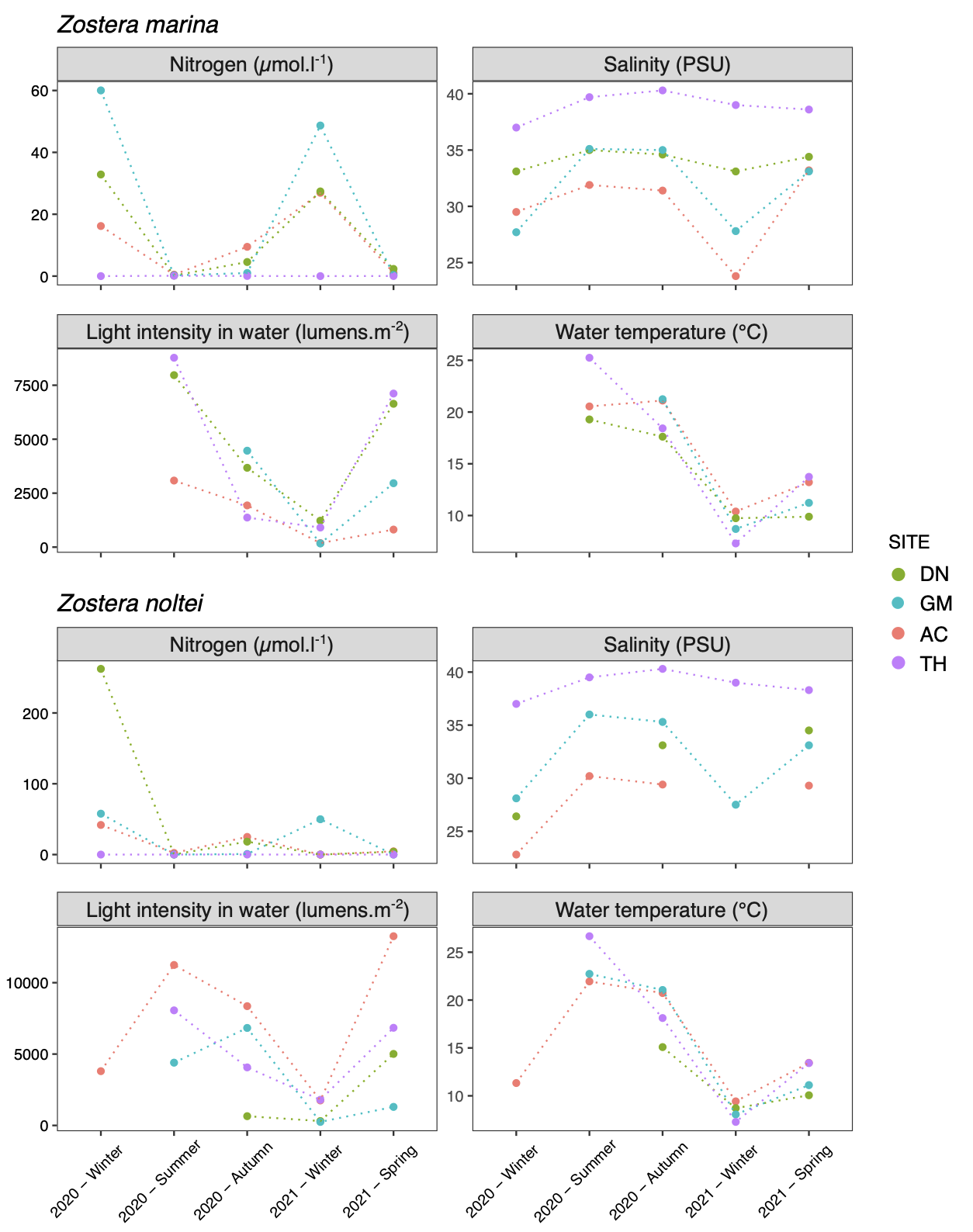
Figure 4 - Environmental conditions measured at each study site in Zostera marina and Z. noltei habitats (DN: Dinard, GM: Golfe du Morbihan, AC: Arcachon, TH: Thau). Light intensity measured during immersion was averaged over the day, excluding the night period. Nitrogen corresponds to ammonium + nitrate + nitrite (dissolved inorganic nitrogen).
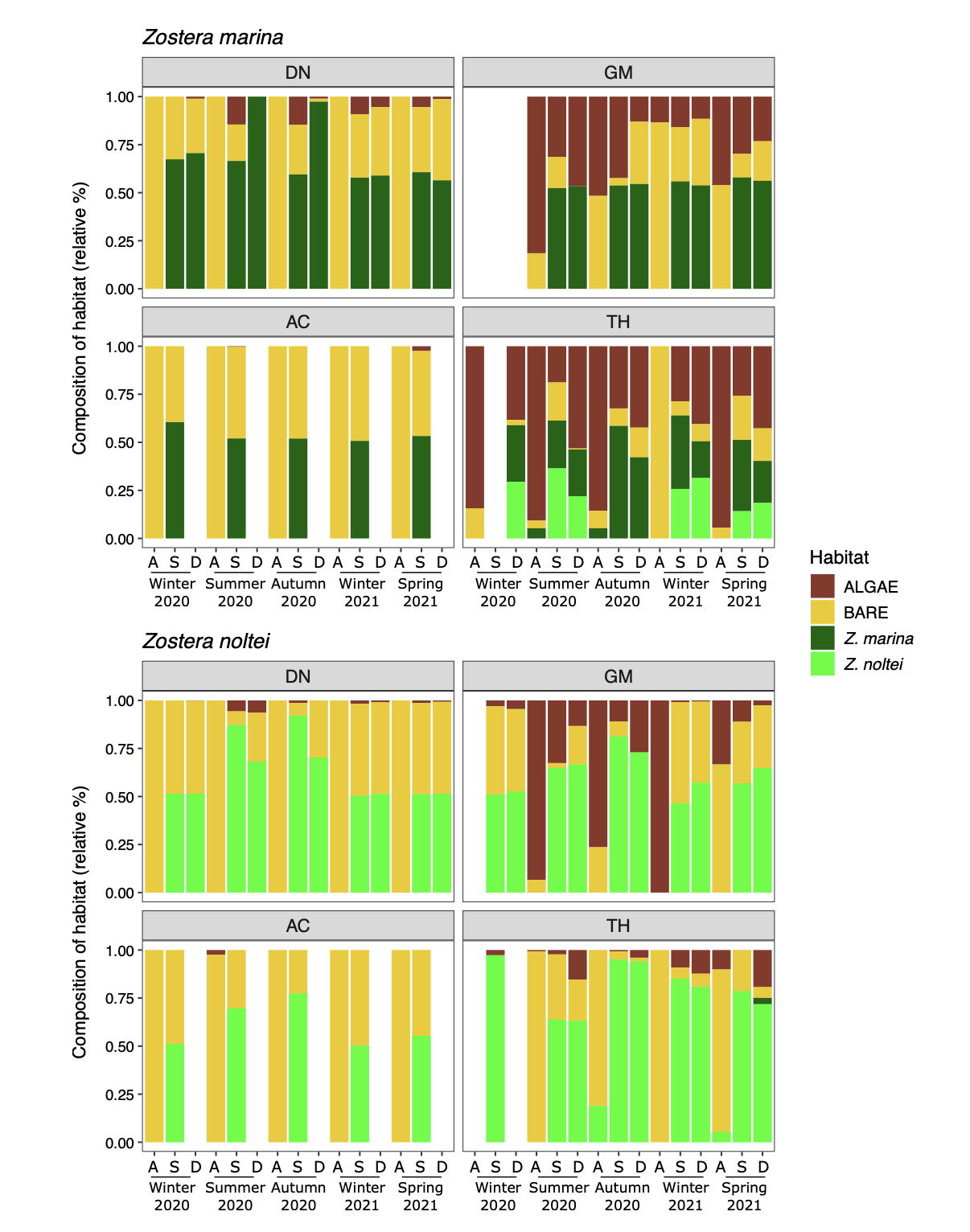
Figure 5 - Seasonal variations of the relative percentage of cover by seagrasses, algae and bare sediment for each combination Species × Site × Modality. Sites: DN: Dinard, GM: Golfe du Morbihan, AC: Arcachon, TH: Thau. Modalities: A: bare sediment, S: stable, D: dynamic. Data are averaged per modality (N=18).
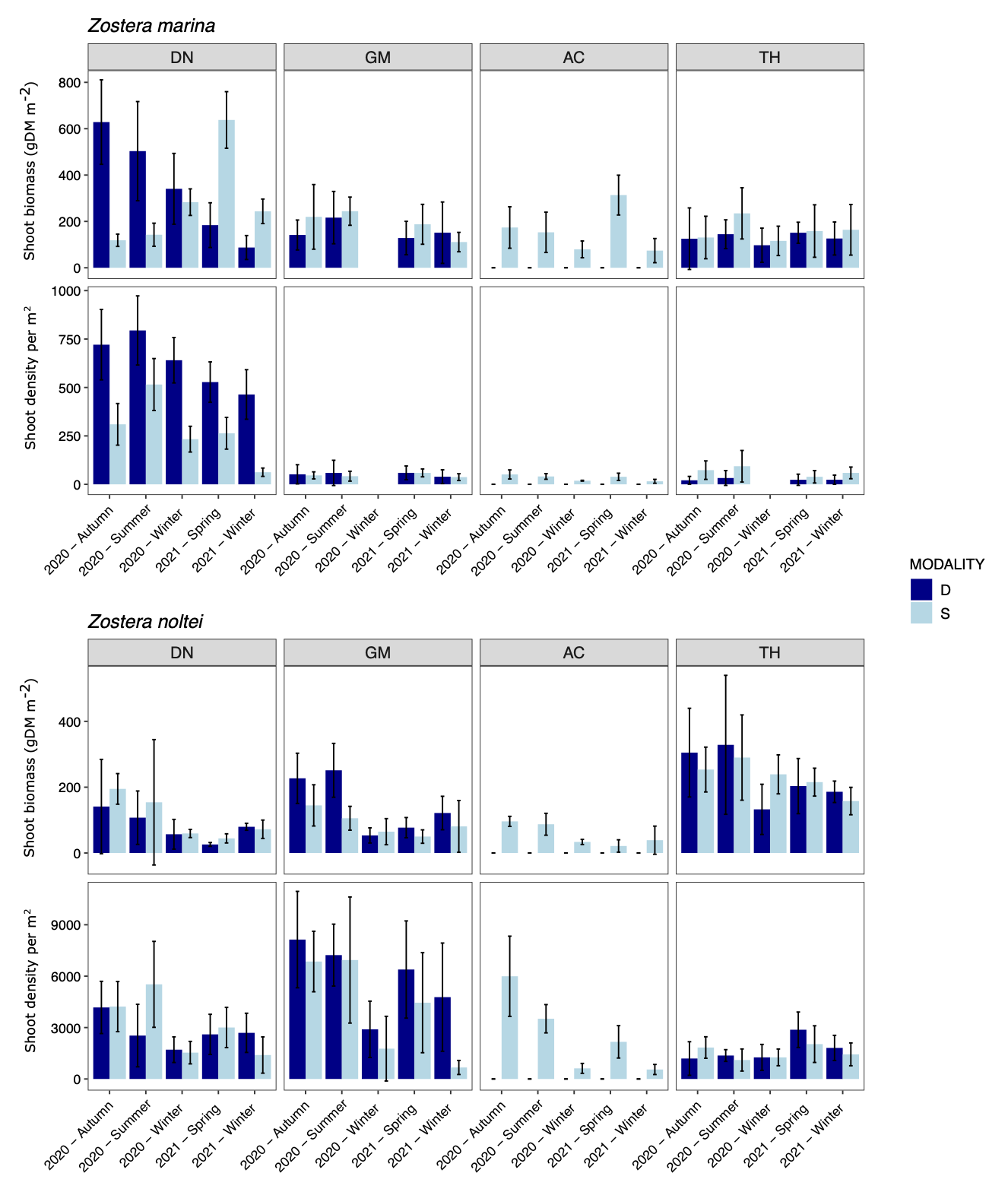
Figure 6 - Shoot biomass per m2 (above ground + belowground dry mass) and shoot density in Z. marina and Z. noltei beds at the four sites (DN: Dinard, GM: Golfe du Morbihan, AC: Arcachon, TH: Thau) and the two modalities (D: dynamic in dark blue and S: stable in light blue) during the study period. Mean ± sd (n=9).
Figure 7 - Epifaunal biomass and density associated with Z. marina and Z. noltei habitats at the four sites and the three modalities during two seasons. Biomass in ash free dry mass per m2. Sites: DN: Dinard, GM: Golfe du Morbihan, AC: Arcachon, TH: Thau. Modalities: A: bare sediment in grey, D: dynamic in dark blue and S: stable in light blue. Mean ± sd (N=3).
Benthic fluxes
The measurement of benthic O2 fluxes was well reproducible, with relative small variations between replicates (Fig. 8). Most values were < 5 mmol O2 h-1 m-2 in both Z. marina and Z. noltei habitat. Respiration and production rarely exceeded 15 mmol O2 h-1 m-2. Season, site and position within the meadow (i.e. modality) induced variability. Data are taken from the “benthic_fluxes” table.
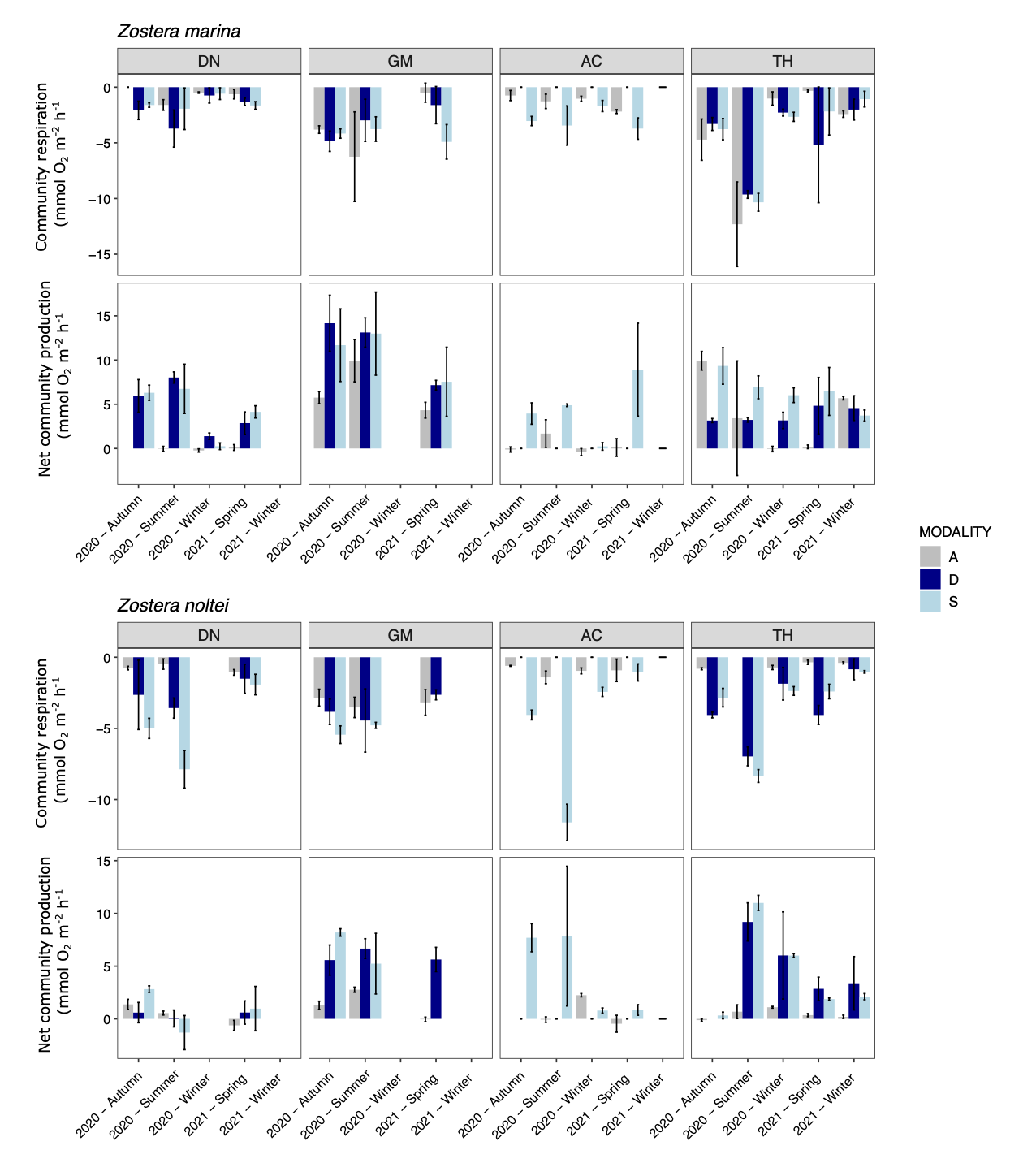
Figure 8 - Seasonal O2 fluxes measured during dark (community respiration) and light (net community production) incubations for the four sites and the three modalities for Z. marina and Z. noltei. A: bare sediment in grey, D: dynamic in dark blue and S: stable in light blue. Mean ± sd (N=3).
Data, scripts, code, and supplementary information availability
Data and supplementary information are available online in Lacoste et al. 2023: https://doi.org/10.17882/97962.
Acknowledgments
This study is part of the Life MarHa project funded by the European Union. The authors thank Benoît Gouillieux, Benjamin Guyonnet, Patrik Le Gall, Françoise Dagault, Titouan Brébant, Aurore Lejolivet, Régis Hocdé, Benoît de Voguier, Laurent Levêque, Ludovic Devaux and Dominique Munaron for their help.
Preprint version 4 of this article has been peer-reviewed and recommended by Peer Community In Ecology (https://doi.org/10.5281/zenodo.10425140; Bornette, 2024).
Funding
The authors declare that they have received no specific funding for this study.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.