
 CC-BY 4.0
CC-BY 4.0
Introduction
From its outset, Ecology was defined as the science of interactions (Haeckel, 1866; Elton, 1927). Initially centered on direct relationships among species, research increasingly emphasized the importance of indirect interactions, highlighting the interplay between herbivores and their predators and how it affected the structure, function and stability of ecosystems [(Paine, 1966, 1969; Estes et al., 2011) and review in (Martin et al., 2020)]. Consequently, while the loss of species or populations across the world’s ecosystems is dire, the loss of species interactions is perhaps even more insidious, because it often goes unnoticed (Janzen, 1974; Soulé et al., 2003; Valiente‐Banuet et al., 2015).
The ecology of fear, connecting behaviour, ecology and evolution – Studies on large herbivores and their predators suggest that, whether predators are present or absent, prey will alter their behaviours in reponse to perceived risk. The non-consumptive effects of predator presence, including human hunters, and its consequences on how prey interact with their habitat, is central to the conceptual framework of "the ecology of fear” (Brown et al., 1999; Zanette & Clinchy, 2020; Potratz et al., 2024), and to the idea of a “landscape of fear” (Laundré et al., 2001; Gaynor et al., 2019, 2021; Zanette & Clinchy, 2020; Palmer et al., 2022). Focusing on ungulates, and deer in particular, Altendorf et al. (2001) predicted that, in presence of predators, deer should spend less time foraging at any given location, limit or avoid using portions of their habitat perceived as more exposed to predation (e.g. because of high visibility), or favour sites perceived as refuges from risk (e.g. dense vegetation) (Williams et al., 2008; Kuijper et al., 2013; Padié et al., 2015; Bonnot et al., 2017; Martin et al., 2018; Clare et al., 2023). Conversely, absence of risk should eliminate the spatial effects of risk on foraging (Zanette & Clinchy, 2020; Wójcicki & Borowski, 2023) and intensify use of the vegetation. Beyond these direct or indirect day-to-day impacts on ecological processes (e.g. Sih et al. 2012), prey behavioural adjustments to the level of risk could also, over time, select for prey traits and behaviours better adapted to the risk level of a given community (Bøhn & Amundsen, 2004; Sih et al., 2004; Réale et al., 2007; Ellers et al., 2012; Estes et al., 2013; Zanette & Clinchy, 2020; Wójcicki & Borowski, 2023).
Questions to address – Despite recent advances, we still need to better understand how animal behaviour, ecology and evolution interact to shape ecological dynamics, behavioural traits, and habitat selection (Réale et al., 2007; Sih et al., 2012; Potratz et al., 2024). Such an understanding would also improve conservation strategies (Kuijper et al., 2016; Martin et al., 2020; Chitwood et al., 2022; Potratz et al., 2024). Here, we use empirical and experimental studies carried out since 1989 on the interplay between deer, vegetation and predation risk, to assess (1) the links between the presence of risk and the effects that deer have on forest ecosystems, (2) whether different metrics of behavioural responses varied in relation to the presence or absence of hunting and natural predators, and (3) how risk affected deer habitat selection.
A life-size laboratory – We took advantage of the introduction, at the end of the 19th century, of Sitka black-tailed deer (Odocoileus hemionus sitkensis) to Haida Gwaii, a remote archipelago in British Columbia, western Canada (Fig. 1) lacking the natural predators these deer are exposed to on the adjacent mainland (Golumbia et al., 2008). The archipelago provided islands with and without deer, and, where deer were present, islands with and without a history of hunting by people. On two islands, severe culling was carried out on a deer population previously without predation risk and where deer browsing had a dramatic impact on vegetation and fauna (Allombert et al. 2005b, 2005a; Stockton et al. 2005; Martin et al. 2010). We complemented our long-term data on Haida Gwaii with data from the Central Coast of British Columbia where deer always faced natural predators in addition to hunters (Darimont & Paquet, 2001, 2002; Darimont et al., 2007). There, black-tailed deer represent over 80% of wolf diet (Darimont & Paquet, 2001).
Fear and vegetation – Based on prior studies, we expected that absence of risk would be associated with severe impact on forest vegetation cover and diversity (Côté et al., 2004; Martin et al., 2010; Stockton et al., 2005), with understory vegetation restricted to a small group of deer-resistant species, even if only represented as severely browsed and stunted individuals (Martin et al., 2010). We also expected that culling, in a deer population initially not exposed to risk, would trigger a positive response in vegetation cover.
Fear and behaviour – We predicted that severe experimental culls and associated changes in perception of risk would affect anti-predator behaviours, possibly years after the culls (Martin & Baltzinger 2002; Gaynor et al. 2019; Clare et al. 2023; Wójcicki & Borowski 2023). In particular we expected to see longer deer flight initiation distances (FID) after detection of a potential human threat, longer distances travelled after detection, a reduction in deer use of foreign bait and a lower likelihood of deer being trapped in the post-culls deer population. We also predicted changes in deer diel activity towards being more nocturnal when compared to populations not exposed to risk from day-hunting humans (Kilgo et al., 1998; Bonnot et al., 2020). Conversely, we predicted that in populations not exposed to risk, lower levels of anti-predator behaviours would be key in explaining the observed severity of deer impact on the vegetation.
Fear and habitat selection – Previous studies have shown that, in some systems, prey favoured foraging in more open habitats because of better visibility of an approaching threat (Gigliotti et al., 2021; Kamaru et al., 2024). In other studies, in particular on deer, elevated risk resulted in prey avoiding open habitats for foraging (Kilgo et al., 1998; Creel et al., 2005; Bonnot et al., 2013; Dellinger et al., 2019). When wolves were introduced to Coronation Island in SE Alaska (Klein, 1995) their extensive use of shorelines came with a dramatic shift by deer away from the coastal portions of the island [for use of shorelines by foraging wolves; see also Darimont & Reimchen (2002) on deer seasonal use of salmon, and Roffler et al. (2023)]. Hunters access to deer on the BC Central Coast, essentially roadless, and on Haida Gwaii, is often by boat [L. Vigneault pers. com. and Irvine & Thorley (2024)]. This, and our observations of deer foraging frequently on seaweeds beached or attached on intertidal shores (Bonnot et al., 2016) led us to predict that the lack of cover in intertidal areas, the limitation of escape routes by the ocean, and wave sounds hindering auditory detection, might decrease shoreline use in presence of risk from predators or hunters. We tested this prediction indirectly by comparing the proportion of marine algae in deer diet under contrasting risk contexts.
Material and Methods
Haida Gwaii and the study sites
Haida Gwaii is characterized by a humid temperate-oceanic climate, with mean annual temperature of 8.5°C (Banner et al., 2014). Most of the archipelago is covered by temperate rainforests dominated by western hemlock (Tsuga heterophylla), western redcedar (Thuja plicata), and Sitka spruce (Picea sitchensis). Open terestrial habitats are restricted to the alpine zones and to extensive bogs on NE Graham Island.
Native to coastal British Columbia, but not to Haida Gwaii, Sitka black-tailed deer were introduced to the archipelago in the late 19th century (Golumbia et al., 2008), colonizing all but a few small islands. The absence of natural predators on the archipelago allowed the deer population to thrive. The occurrence of a few reference islands that never supported deer made it possible to demonstrate that, on islands with long-term deer presence, independent of island size, deer herbivory was the main factor structuring plant and animal communities (Martin & Baltzinger 2002, Gaston et al. 2006, Martin et al. 2010, Chollet et al. 2013), with important consequences on belowground processes (Chollet, Maillard, et al., 2021; Maillard et al., 2021). Recurrent experimental culls on two islands allowed us to monitor the response of the aboveground vegetation and avifauna for 13 years (Chollet et al. 2016). These results accumulated over the different phases of the long-term RGIS project https://rgis.cefe.cnrs.fr.
The three islands in Laskeek Bay (52°53'12"N, 131°35'20"W) where we focused our study on deer behaviour (Reef 249 ha, Kunga 395 ha, and East Limestone Island 48 ha) (Table 1, Fig. 1) have all had deer present for over 60 years at the time of study (Vila, Guibal, et al., 2004; Vila, Torre, et al., 2004) and had no history of hunting. Covered by mature forests without human settlements or activities, these islands were characterized by closed canopy forests with open species-poor understories (Martin et al. 1995, Stockton et al. 2005, Martin et al. 2010) representative of severe deer impacts that we documented at the scale of the archipelago (Martin et al. 2010; Chollet et al. 2015, 2021a). Rocky shorelines and areas exposed at low tide fringe the islands. On most islands, deer density was estimated around 30 deer / km² (Daufresne & Martin, 1997; Martin et al., 2010; Stockton et al., 2005) an estimate that has been repeatedly confirmed in the course of deer culls (Gaston, Golumbia, et al., 2008; Irvine & Thorley, 2024).
Table 1 - List of islands included in the study and their key characteristics. Risk hist. = Risk history; Deer: Y = present, N = absent; Island = island name; Hunting: Y = seasonal hunting, Y(cull) = repeated culls; N = no hunting; Predators: Y = present (wolves and possibly cougar and grizzly bear), N = absent; Veg. plots: figures = number of standardized plots on vegetation structure and composition, N = no standardized sampling; Behav.: Y = all behavioural data collected, N = none collected; Isotope = collection of bones for the stable isotope data for the study on the use of shorelines by deer: Y = collected and analyzed, N = none collected; Reef <1997 = conditions on Reef prior to experimental culls; Reef >1997 = conditions after the culls that occurred between 1997 and 2008; * = Islands that only provided stable isotope data for the study on shoreline use. + = bones were sampled in the area of BC Central Coast between Yeo and Bella Bella.
Risk hist. | Deer | Island | Region | Area | Hunting | Hunting + Predators | Veg. plots | Behav. | Isotope |
|---|---|---|---|---|---|---|---|---|---|
No-risk | N | Low | Haida Gwaii | 9 ha | N | N | 5 | N | N |
No-risk | N | South-Low | Haida Gwaii | 13 ha | N | N | 5 | N | N |
No-risk | N | Lost | Haida Gwaii | 5 ha | N | N | 5 | N | N |
No-risk | Y | East Limestone | Haida Gwaii | 48 ha | N | N | 10 | Y | Y |
No-risk | Y | Kunga | Haida Gwaii | 395 ha | N | N | 20 | Y | Y |
No-risk | Y | Faraday* | Haida Gwaii | 308 ha | N | N | N | N | Y |
No-risk | Y | Murchison* | Haida Gwaii | 425 ha | N | N | N | N | Y |
No-risk | Y | Reef <1997 | Haida Gwaii | 249 ha | N | N | 22 | N | Y |
Risk | Y | Reef >1997 | Haida Gwaii | 249 ha | Y (cull) | N | 22 | Y | Y |
Risk | Y | Graham | Haida Gwaii | 6361 km² | Y | N | 20 | N | Y |
Risk | Y | Yeo | Coastal BC | 95 km² | Y | Y | 7 | N | Y+ |
Reef Island, devoid of hunting prior to 1997, was subjected to repeated and severe deer culls between 1997 and 2008 (Chollet et al. 2016). From September 1997 to February 1999, over 80% of the initial deer population was culled (Gaston, Sharpe, et al., 2008). Recent archipelago-wide genetic analyses confirmed that these culls caused a severe population bottleneck and that the current deer population of Reef Island consists of descendants from the handful of animals left after the culls (Burgess et al., 2022a; b, 2023). As local deer survival is typically less than 10 years (JLM et al. unpubl.), most or all individuals involved in our study during 2011-2014 must have been born after the initial culls. In 2014, Reef Island had a deer population of about 15 deer / km², thus an estimated 30 to 40 deer present, representing about half of the pre-cull population, and a partially, but dramatically, recovered understory vegetation (Chollet et al. 2016).
We also studied a portion of Graham Island (6,361 km²), the largest island of the Haida Gwaii archipelago, where deer have been widespread since the early 20th century (Golumbia et al., 2008) and have been exposed to human hunting ever since. Graham is also home to a population of black bear (Ursus americanus) that sometimes prey on deer fawns (Mathews & Porter, 1988; Ballard et al., 2001). Deer densities on Graham have been estimated to exceed 13 deer / km² (Engelstoft, 2001; Engelstoft et al., 2008). Its forests are characterized by low vegetation cover in the understory [this study and (Chollet, Baltzinger, et al., 2021)].
Finally, we complemented the sites selected on Haida Gwaii with data on the vegetation from Yeo Island (Table 1, Fig.1), situated about 15 km north of Bella Bella, close to the mainland of British Columbia. Deer are native to Yeo and exposed there to predators such as wolves (Canis lupus) (Darimont et al., 2007), black and brown (Ursus arctos) bears, and cougars (Felis concolor), and to human hunting. Forestry operations occur in parts of Yeo and Graham islands but we restricted our investigations to unlogged mature forests. We lack reliable estimates of current deer densities for the coastal region. However, a modelling exercise analysing the relationships between habitat, deer and wolves posited that, in the long term, a dense understory would be compatible with high deer densities exposed to predation by wolves (Kirchhoff & Person, 2008).
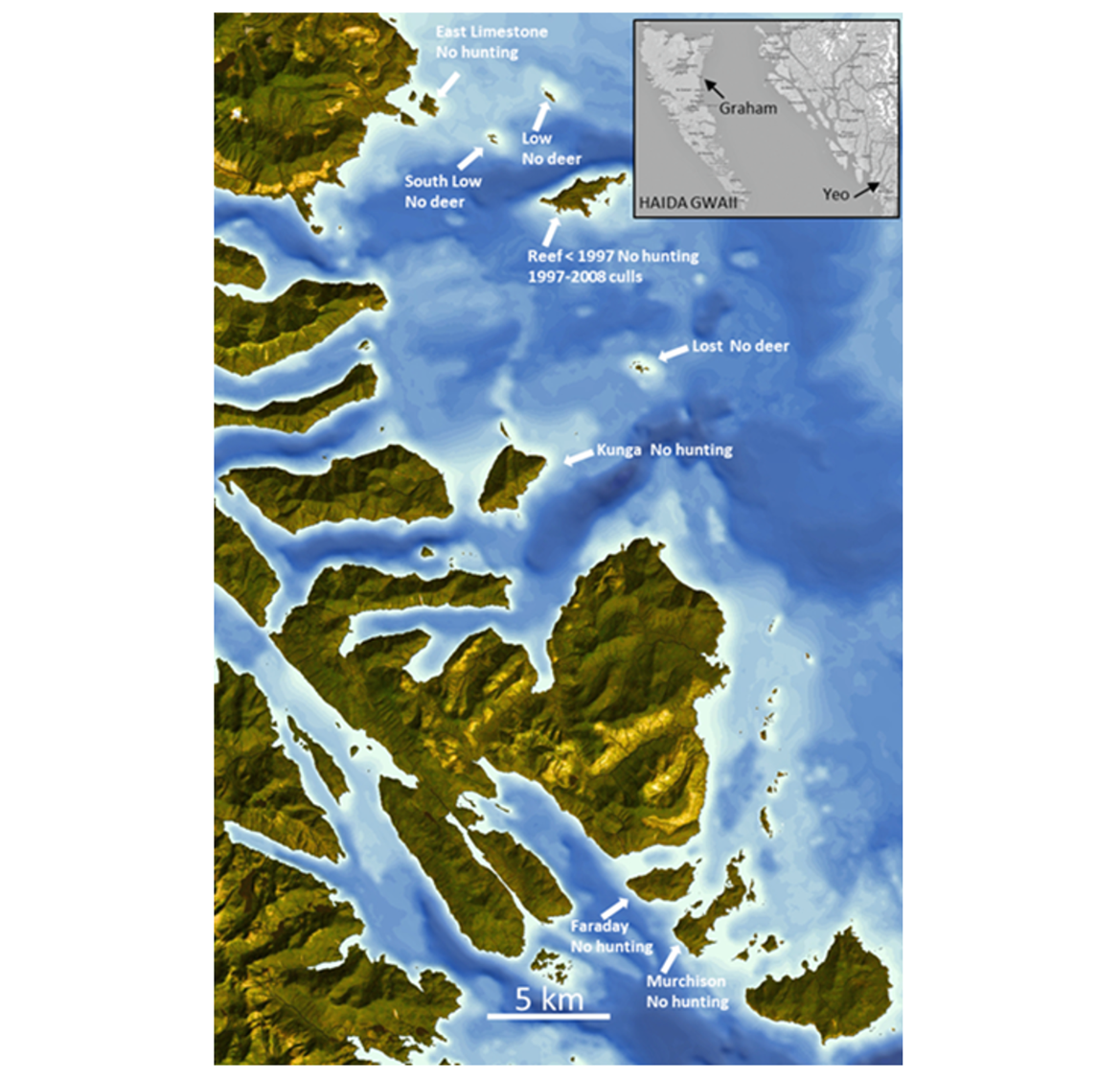
Figure 1 - Map of the study area. Inset = localisation of Haida Gwaii and of Graham and Yeo islands in western British Columbia, Canada. Main map = location of the islands studied in the central east coast of Haida Gwaii, BC, Canada. Yeo = hunting and natural predators. © of map Gowgaia Institute.
We thus had access to study sites varying in history of deer presence, in hunting history and in deer exposure to large carnivores. On East Limestone, Kunga and Reef we also had access to 27, 23 and 8 marked deer, respectively, that we ear-tagged and GPS collared during a project that took place from 2011 to 2013.
Does vegetation cover and diversity vary with predation risk?
To assess the links between the vegetation and predation risk, we compared the understory vegetation among sites with different risk histories (Table 1). We used 3.6 m radius (50 m²) vegetation plots (Table 1) to estimate the % cover of plant species in the 0 to 1.5 m vegetation layer directly accessible to deer browsing for all islands, except Murchison and Faraday. Details for Graham Island sites can be found in Chollet et al. (2021a) and for the other islands from Haida Gwaii in Stockton et al. (2005). We grouped the plant species data into six groups: young conifers, young deciduous trees, shrubs, ferns, forbs, and other herbaceous plants. We used a Principal Component Analysis (PCA) on centered and standardized cover data in R (R-4.1.0) (Racine, 2012) [prcomp function in R (Team, 2018)] to characterize variation in the cover of these plant groups in the understory when deer are present or not and under different contexts of risk from hunters and/or predators.
Does deer fear vary with predation risk?
To assess, quantify and compare deer behaviour in relation to risk history we studied deer Flight Initiation Distance (FID), Distance travelled during flight (Dtravel), and deer response to bait and traps in the different deer populations. We also compared deer diel activity patterns in relation to risk by using activity data collected by automatic cameras.
FID experiments
FID (Flight Initiation Distance) is the distance at which an animal starts moving away at the approach of what it perceives as a potential threat (Ydenberg & Dill, 1986). FID has been considered as a key method to disentangle the “economics” of anti-predatory behaviour, as flight occurs where the decreasing value of remaining, and the increasing cost of not fleeing, intercept (Cooper Jr, 2008). According to Lima & Dill (1990) FID should, under equal resource level, be shorter in safe areas, and longer in risky areas. We therefore used FID as a metric of wariness, using the following protocol: once the observer detected an individual, it was identified by means of its ear tag number or its morphological features. Then the observer walked slowly and calmly (~2 km/hr) towards the deer, avoiding eye contact. The observer stopped walking when the deer changed its initial behaviour and started moving away, but continued observing the animal. We recorded and used as focal variables two distances: distance from the observer at which the deer starts to move away (FID), and Distance travelled (Dtravel) by the deer between its first location (when flight began) and its second location where it stopped moving away and resumed (foraging) activity. We measured these distances by footstep lengths calibrated in meters after each encounter. For deer native to Reef, FID data included deer moving out of sight from the observer. In these four instances we recorded the distance to where it disappeared. This led to a conservative estimate of Dtravel for such cases. To avoid the confounding effects of pseudo-replication and deer habituation to the experiment, we discarded repeated measurements on the same individual and only considered the first FID experiment done in a given sequence.
Data on FID was obtained on islands without risk (Kunga and East Limestone), and on an island intially without risk but subjected to culling (Post-culls Reef). As the expression of fear can be affected by resource availability (Cooper 2008) we created a translocation experiment by moving six adult does from Kunga island, where they were not exposed to hunting and where the understory was heavily browsed, to Reef Island, where heavy culling in the 1990s had created a much denser understory. On Kunga, the translocated does had been box-trapped, marked and collared as adults in 2011 and had been recaptured multiple times (5 to 21 times each) (Le Saout, Chollet, et al., 2014; Bonnot et al., 2016). Three of the six translocated animals had been subjected to five, seven and 19 FID measures on Kunga in 2011 and 2012. We replaced their GPS collars before their translocation to Reef in September 2013 where these animals settled among the local deer born after the culls [(Burgess et al., 2022a, 2023) and M.A. Russello pers. com.]. In spring 2014, five of the translocated animals were still present on Reef. We tested FID behaviours of translocated and local deer in the same way as on Kunga.
We analysed the two distance variables for these deer samples with a linear model (lm function in R) to fit the linear regression model with the distance variable (FID or Dtravel, both log10 transformed to ensure normality) used as a response variable to compare treatments. We also analysed, when feasible within deer samples, the effect of deer status (marked/unmarked), sex (M/F) and age class (adult, young) as well as effect of year on FID and Dtravel. We used pairwise comparisons of the linear models with the emmeans package in R which provided t-tests to compare the effects of these variables on our distance variables within deer samples.
Response to bait and traps
Neophobia, an adverse reaction to novelty, is an important trait that allows animals to minimize exposure to threats (Greenberg & Mettke-Hofmann, 2001; Monestier et al., 2017). Between 2011-2014, we looked at deer use of stations baited with apples and quantified interest in this foreign food by using camera traps (RECONYX PC900) (Le Saout et al., 2015). We also used traps baited with apples to assess the propensity to get trapped as a proxy for exploration behaviour in the presence of a foreign object. We baited each bait station daily with 1.5 to 2 apples unless weather impeded fieldwork. For trapping we placed one chopped apple outside the trap entrance, and another one at the furthest end inside the trap near the trigger. Depending on the requirements of each stage of the study, the cameras at bait stations were programmed to acquire from ten to 99 pictures every time the motion sensors were triggered, with a 1s intervals between pictures. A built-in infrared flash with no red glow allowed us to capture images at night or under low light conditions. The following data were recorded: whether or not the deer had been trapped (unmarked/marked); bait presence at the station (in case consumed by previous visitors) (yes/no); bait consumption if bait present (yes/no); the time at the beginning and end of a sequence. We also recorded if the deer investigated the ground area where the bait had been before (if bait was absent, yes/no). In 2011 bait stations were monitored with automatic cameras for 15 days on East Limestone (no-risk, 4 locations), 12 days on Kunga (no-risk, 4 locations) and 28 days on Reef (post-culls, 8 locations).
In the context of the translocation experiment we applied similar protocols on Reef in the spring of 2014, using six bait stations distributed across five locations, and keeping them active for fourteen consecutive days. We compared responses to bait and traps on Reef Island with the past behaviour of these deer on Kunga and with the behaviour of the deer resident on Reef. No hunting took place on Reef Island during the eight months the translocated animals spent there from September 2013 to May 2014.
We recorded the tag identity of the marked individuals and identified unmarked individuals using physical features such as antler shape, fur marks, scars etc. We used the recorded start time and end time of a feeding sequence to compute the time a deer spent at a bait station when bait was present. We considered time spent at a bait station with bait as a measure of bait friendliness (the willingness to check and consume bait) as in Chamaillé-Jammes et al. (2014). We used an analysis of variance on log10 transformed minutes (aov and emmeans functions in R) to compare “time-spent” among treatments and between marked and unmarked deer within a treatment. We analysed in the same way time spent at bait stations where bait had been consumed during previous visits. We also compared trapping rates between categories using a Welch two sample t-test in R and analysed the propensity of individual deer to be trapped repeatedly.
Daily activity rythms
To assess diel activity (e.g., if risk from daylight hunters led to more nocturnal feeding) we used the time recorded by automatic cameras in all deer observations collected during a study on deer vigilance (Le Saout et al., 2015), and during our investigation on the use of bait stations. We used the time recorded at the outset of each observation to assign observations to day or night. We defined day as the period between civil twilight start (morning) and end (evening) for that date, using Reef Island as the reference locality for civil twilights (time difference with the two other localities is < 20s). (https://www.timeanddate.com/sun/@6118904?month=5&year=2011). We compared the proportion of day and night observations among sites with a Pearson’s Chi-squared test (R function chisq.test).
Does shoreline use by deer vary with risk or amount of understory vegetation?
Because marine plants are enriched in 13C compared to terrestrial plants (Balasse et al., 2005; Richards et al., 2006; Schulting et al., 2008), the isotopic signatures of tissues from coastal herbivores can be used to estimate the relative contribution of terrestrial versus marine plants in their diet (DeNiro & Epstein, 1978). Stable isotope ratios are expressed in the standard delta (δ) notation, defined as parts per mil (‰) deviation from a standard:
δX=[(R in sample/R in reference) -1]*1000 (‰)
δX gives the deviation between the samples’ isotopic ratio and the ratio obtained from an international standard, “R in sample” is the isotopic ratio considered, in our case 13C/12C and 15N/14N. For Carbon, “R in reference” refers to the standard Pee Dee Belemnite (PDB).
δ15N typically gets enriched by about 3‰ when going from one trophic level to the next. At the consumer level the whole animal body δ13C closely reflects its concentration in the diet but its fractioning varies among tissues. Values of concentration gain vary from + 1.5‰ in muscle, to + 5‰ in consumer’s collagen tissue when compared to whole body values (DeNiro & Epstein, 1978, 1981; Bocherens, 1999; Kelly, 2000; Bocherens & Drucker, 2003; Camin et al., 2016).
To test our prediction that risk from hunters and/or predators could limit the use of the intertidal areas for foraging, we analysed the proportion of stable isotopes of terrestrial plant or seaweed origin in deer diets on different islands, using deer jaw bones from all the study populations, to which we added bone samples available from Murchison and Faraday Islands, both hunting-free islands (Fig.1 and Table 1). We obtained these bones either from hunting, or from deer that had died from natural causes. On the larger islands (Kunga, Graham and the coastal area adjacent to Yeo) we restricted bone collecting whether through hunting or other means to less than 1000 m from the shoreline. We obtained 10 jaw bones from East Limestone, 12 from Kunga, 10 from Murchison, six from Faraday, seven from pre-cull Reef, all without hunting, six from Post-culls Reef, collected 10 years or more after the initial culls (2008 to 2014), 23 from Graham Island (hunting), and 17 from the central coast district between Bella Bella and Yeo Island (hunting + predators). We cleaned and dried bones in the field.
For comparison of isotopic composition, we also collected seaweed and plant material across sites. For seaweeds we collected or had access to samples from one to thirty individuals of 41 seaweed species (19 species of brown, 5 green, and 19 red) collected on Reef Island, Louise Island, and Kunghit Island on Haida Gwaii, and on the coastal mainland. We preserved them according to the protocols of Salomon et al. (2008). We also collected samples of 23 dominant plant species in Laskeek Bay (Reef and East Limestone islands) and on the coastal mainland, including 4 species of coniferous trees, 2 deciduous trees, 9 deciduous shrubs, 1 evergreen shrub, 5 ferns and 5 grasses. We rinsed fresh samples in distilled water to avoid contamination, pre-dried them in the field-camp cabin and fully dried them in the lab at 40°C for 24 hours.
We ground dry bone and plant samples to a granulometry of less then 0.7mm. We followed protocols defined by late F. Catzeflis based on DeNiro & Epstein (1981), Bocherens et al. (1988), and Bocherens, Hervé et al. (1991) to extract bone collagen from bone powder in the laboratory. We analysed the isotopic composition of the plant and collagen samples on CO2 and N2 obtained by sample combustion and analysed on a mass spectrometer. On the basis of isotopic similarities, plants, except conifers and the evergreen shrub Salal (Gaultheria shallon), were combined in one group (DecHerbs). All seaweeds had a similar, restricted spread of isotopic signatures and we pooled them into a single group (Seaweed).
We analysed the isotopic ratios obtained for our samples using a Bayesian multiple source mixing model (MixSIAR package in R) (Stock, Jackson, et al., 2018; Stock, Semmens, et al., 2018) which estimates the proportions of source contributions (here terrestrial and marine plants) to a mixture (bones used as a proxy for deer diet) (Bocherens and Drucker 2003).
Results
Understory vegetation varied with risk history
High vegetation cover and diversity was found on islands without deer and on those where deer were present but exposed to predators and hunting (Figs. 2 and 3). Their understories were dominated by forbs, shrubs, and ferns (Figs. 2 and 3). The samples from Reef Island (Post-culls) showed an increase in cover and diversity after the culls, resulting in a large spread of plots. Most had high vegetation cover in the understory (positive scores on PC1 and/or negative scores on PC2), but their cover was dominated by conifer regeneration and grasses and some cover of ferns and shrubs (negative scores on PC2 and low positive scores on PC1) (Figs. 2 and 3). Forests with deer and no hunting (Reef Island prior to culls, Kunga and East Limestone), showed a stark contrast with those from islands without deer or those where deer coexisted with hunters and predators (small narrow ellipse along PC2, Fig. 2). Most plots had very low plant cover and conifers were the only plant group with significant cover (Fig. 3): plots clustered around low cover values for all understory plant groups (low negative scores on PC1 and low positive scores on PC2, Fig. 2). Only three plots from pre-cull Reef and eight from the other islands without hunting had high plant cover (negative scores along PC2, Fig. 2) consisting of conifer regeneration (spruce and hemlock) (Fig. 3). The samples from Graham Island, where yearly moderate hunting has occurred for over a century, were also dominated by plots with low plant cover in the understory (Fig. 2) but had, in adition to conifer cover, more shrub cover and measurably more cover of ferns and forbs in their understory than plots from the smaller islands without hunting (Fig. 3), hence their higher spread along PC1 (Fig. 2).
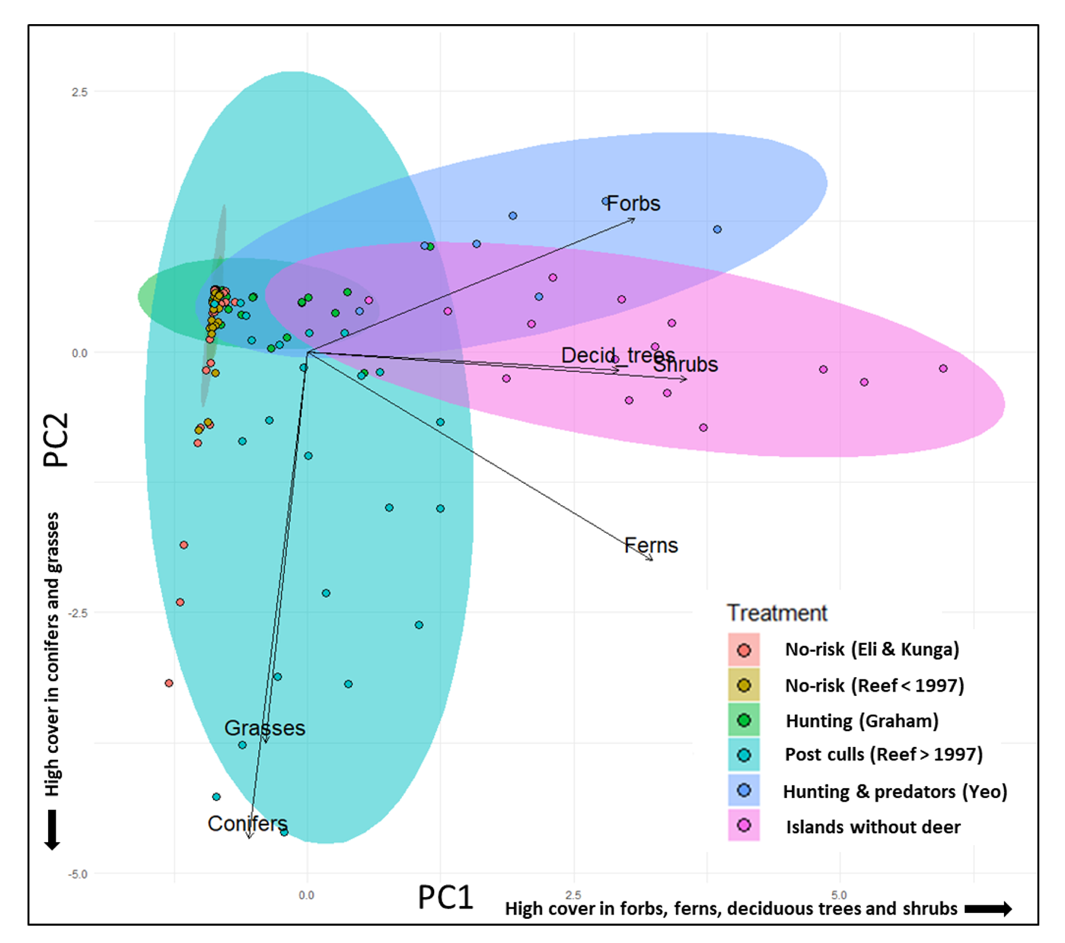
Figure 2 - Scores of the vegetation plots on the two first components of the Principal Component Analysis on understory vegetation cover below 1,5m. PC1 and PC2 summarize 37 and 19% respectively of the variance. Treatments were color coded post-analysis: orange = No-risk East Limestone and Kunga island samples, kahki = No-risk sample from Reef island before the cull (<1997), green = long history of yearly hunting (Graham island); green-blue = risk in the recent past through culls (Reef >1997), blue = hunting and deer predators (Yeo island), pink = reference islands without deer (Low, South Low and Lost islands). The dots refer to the actual coordinates of each plot. The ellipses are the most parsimonious graphical interpretation of the spread of plots in each sample.
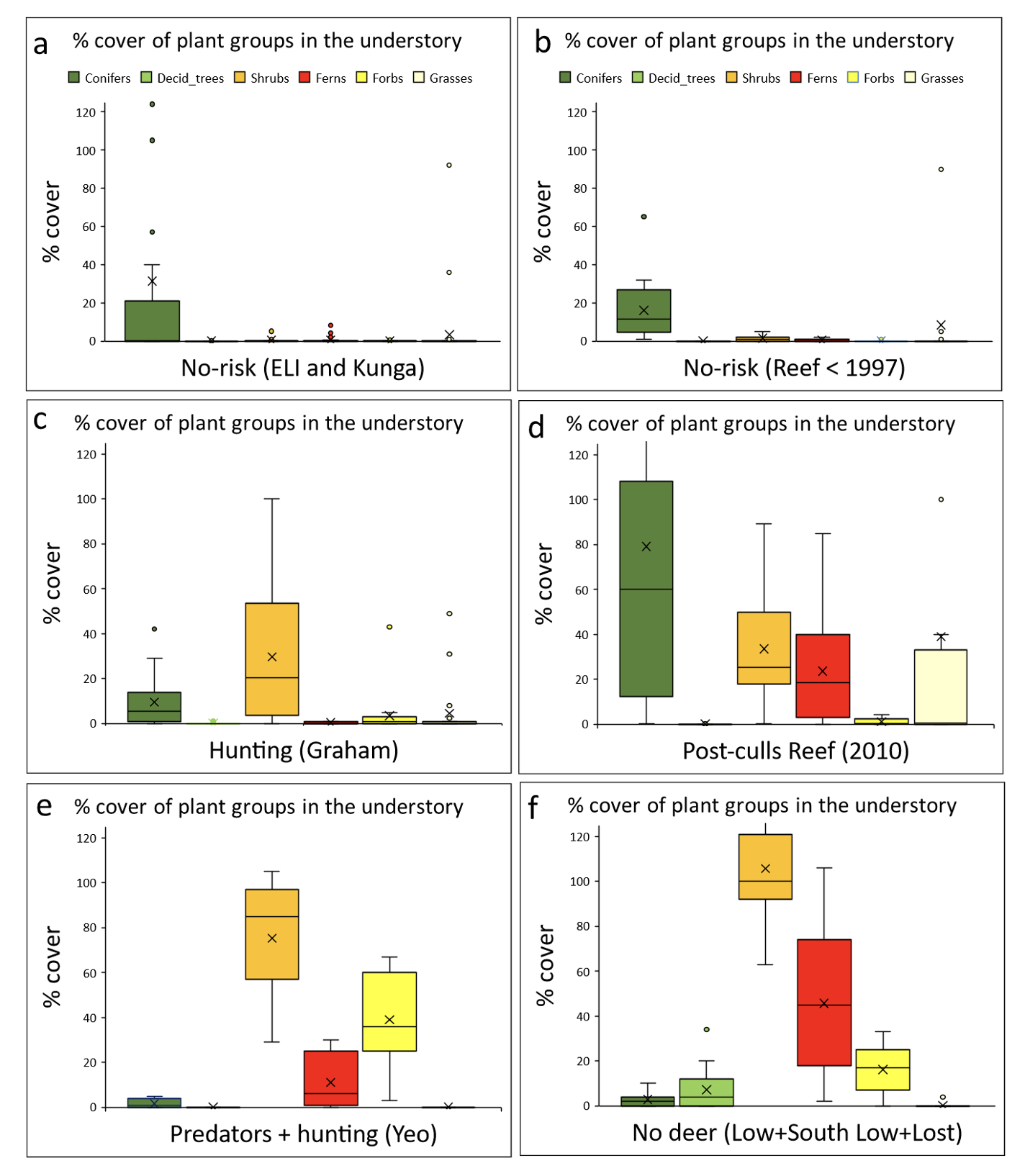
Figure 3 - Percent cover recorded in the 0 to 1.5 m strata for the main understory plant categories in the different treatment categories. The vertical scale has been limited to 125% at the expense of a small number of outliers for conifers on graph 3d, and shrubs on graph 3f. Dark green = conifers, light green = deciduous trees, orange = shrubs, red = ferns, bright yellow = forbs, light yellow = grasses.
Deer fear varied with predation risk
Flight initiation distance
We ran FID assessments on the seven deer treatments during four field seasons from spring 2011 to spring 2014. Most took place in 2011-2012. We retained 218 FID events: 73 from Kunga, 133 from East Limestone, both without risk, and 12 from Post-culls Reef. We had no FID data for Graham and Yeo or for Reef prior to 1997. We recorded 19 FID observations for the Kunga deer eight months after their translocation to Reef Island.
Values did not differ significantly among years within samples (p = 0.60). Nor did sex or age have any effect (p = 0.30, p = 0.62 respectively). Thus we combined sexes, ages and years.
FID varied significantly among treatments [R2 = 0.38, F(3, 233) = 49.28, p < 0.001] (Fig. 4a). FID was lowest for islands without hunting and highest for Post-culls Reef. Among the islands without hunting, FID was lowest on East Limestone and highest for Kunga deer translocated to Reef (Fig. 4a).
Samples from No-risk East Limestone and from Post-culls Reef included marked and unmarked deer. Differences in FID between marked and unmarked deer were non-significant [for East Limestone t-ratio = 1.85, p = 0.07 (6.9 versus 5.5 m); for Reef Island t-ratio = 0.95, p = 0.34 (36 versus 23.6 m)].
Distance travelled
Dtravel varied significantly among treatments [R2 = 0.18, F(3, 229) = 18.04, p < 0.001]. Deer from populations without hunting travelled significantly shorter distances than deer on Reef after the culls (Fig. 4b). Dtravel on East Limestone Island was significantly shorter than on Kunga Island (p = 0.01), and much shorter than on Post-culls Reef Island (Fig. 4b; p < 0.001). Dtravel distances for Kunga deer translocated on Reef did not differ from those on Kunga (p = 0.96), or from Dtravel observed on East Limestone (p = 0.12) and was only one third that of native Reef deer (p < 0.001) (Fig. 4b). On East Limestone, marked deer moved significantly further than unmarked deer [t-ratio = 2.20, p = 0.03, (6.7 m versus 4.9)] but this did not apply to marked and unmarked deer on Reef [t-ratio = 0.17, p = 0.86 (41m versus 36.7)].
Response to bait
Risk category had a significant effect on time spent at bait stations when bait was present (log10 transformed): deer on islands without hunting and deer from Kunga translocated to Reef, spent significantly more time at bait stations with bait than native Reef deer (R2 = 0.14, F(3, 79) = 4.29, p = 0.007; Fig. 4c) and visited them five times more often (0.97 against 0.20; Fig. 4c, Table 2). Bait consumption per visit was three times higher on Kunga and East Limestone than for native deer on Reef (75% against 20%) (Table 2). On East Limestone and Kunga marked deer consumed bait in 50-80% of their visits in contrast to native deer on Reef, where only three of the nine deer trapped and marked consumed bait at bait stations. None of the 14 unmarked deer observed on Reef touched bait which they investigated briefly or ignored altogether (mean visit length 24 s). Length of median values in time spent on a station, less affected by outliers, exceeded 3 min for deer on Kunga and East Limestone but was less than 30 s for native Reef deer.
At Kunga, marked deer spent significantly more time at a station than unmarked deer (7.8 min on average against 5.6 minutes) (t-ratio of contrast estimate = 4.64, p < 0.001). Some unmarked animals visited a bait station without eating bait (mean visit length 54 s). Among deer native to Reef, marked individuals also spent longer at bait stations (t-ratio of contrast estimate = 3.00, p = 0.004) and were the only deer consuming bait (mean visit length 3.0 minutes).
In absence of bait because of previous consumption, visit length at a station was short (Fig. 4d versus Fig. 4c). Visits were longest on Kunga through the effect of outliers spending long periods at sites that had bait recently (Fig. 4d). Time spent at a station never exceeded 4 minutes and was spent sniffing at where bait had been present. On Reef, marked native deer spent significantly longer at stations without bait than unmarked native deer (means of 2.8min versus 0.13 respectively) (t-ratio of contrast estimate = 4.0, p < 0.001). On No-risk Kunga, there was no significant difference between marked and unmarked deer in time spent at stations without bait (84 versus 90 s) (t-ratio of contrast estimate = 1.42, p = 0.16).
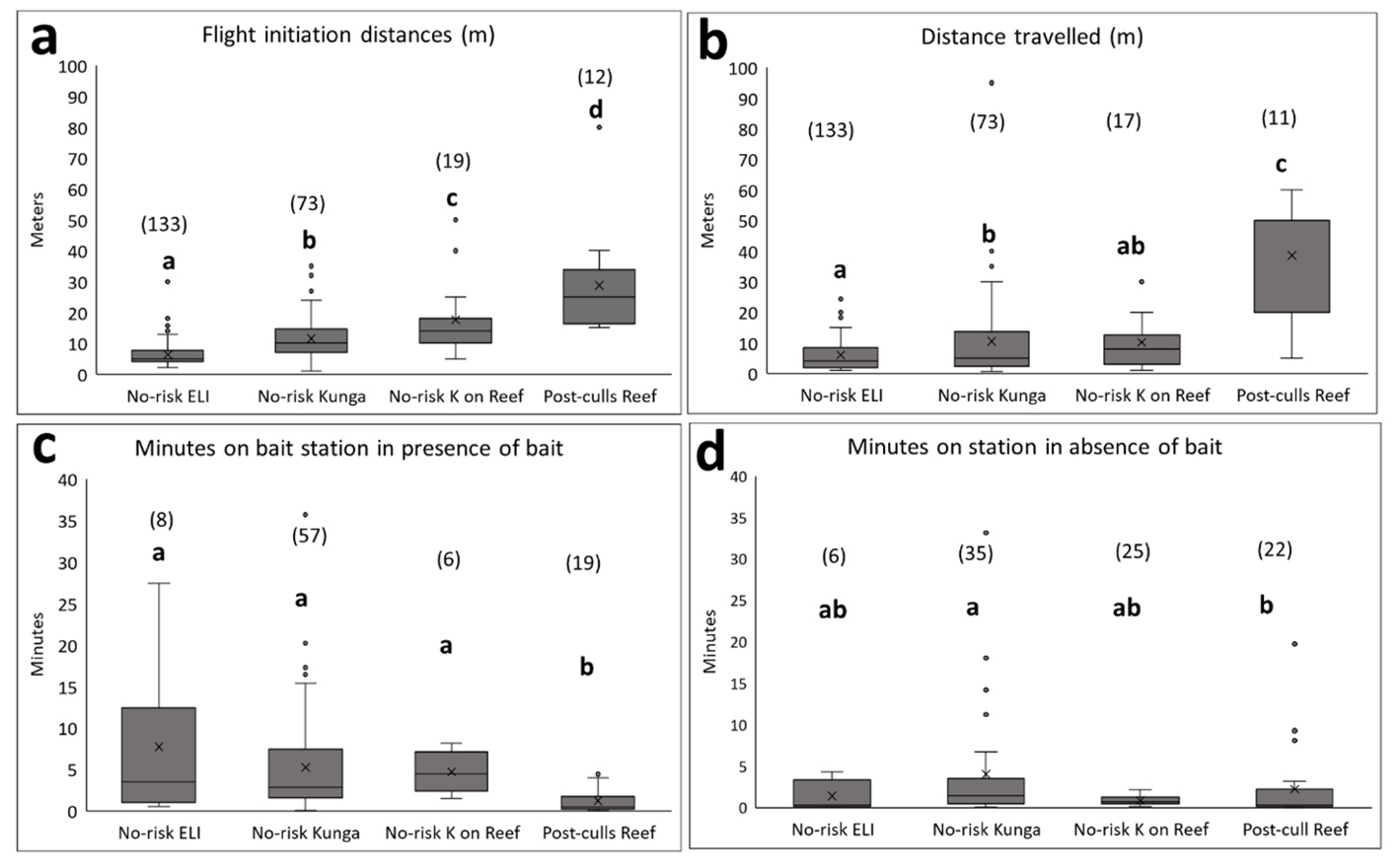
Figure 4 - Boxplots on Flight and travel distance and on time spent at bait stations with bait present or absent. No-risk ELI = East Limestone deer, No-risk Kunga = Kunga deer on Kunga Island, No-risk K on Reef = Kunga deer after translocation to Reef Island, Post-culls Reef = Reef Island deer born into a population that survived the 1997 to 2008 culls. X in box = mean value, solid line in box = median value. Letters indicate results of post-hoc tests on the linear regression model on FID (log10 transformed) as a function of deer categories. Numbers between parentheses = sample sizes.
a - Flight Initiation Distance (FID). All pairwise comparisons were significant: most had p-values < 0.001 except No-risk Kunga – No-risk K on Past culls Reef (p = 0.02) and No-risk K on Post-culls Reef – Post-culls Reef (p = 0.03).
b - Distance travelled after flight initiation (Dtravel). For Post-culls Reef median = 50 m. All pairwise comparisons were significant except for No-risk ELI – No-risk K on Reef (p = 0.12) and No-risk Kunga – No-risk K on Post-culls Reef (p = 0.96). For the pairwise comparisons that were significant all p-values were < 0.001 except No-risk ELI – No-risk Kunga (p = 0.01).
c - Time spent at bait stations with bait present. All comparisons among No-risk categories were non-significant (p-values > 0.80). For the pairwise comparisons that were significant, all p-values were < 0.05.
d - Time spent at bait stations by deer when bait was missing (consumed since station was re-provisioned). All comparisons among No-risk categories were non-significant (p-values > 0.50), except for No-risk Kunga – No-risk K on Post-culls Reef (p = 0.37). Only for the No-risk Kunga – Post-culls Reef comparison did Kunga deer spent significantly more time at stations with no bait left than did deer native to Post-cull Reef (p = 0.04).
Table 2 - Deer visits and use of bait stations with bait present in relation to island/hunting history category. Stations (days) = number of stations set up and number of days each was active; Visits = total number of photographic sequences a deer was captured on camera at a bait station; With bait = number of visits when bait was present; Eat = number of visits in which bait was consumed when present; Mean length (median) = mean value of a visit duration in minutes and the corresponding median value; Without bait = number of visits when bait was absent (= consumed during visits that followed the re-provisioning of the station); Deer = total number of different individuals involved in the experiment; Marked = number of marked individuals among the total number of different individuals involved.
Island/hunting history | Stations (days) | Visits | With bait | Eat | Mean duration (median) | No bait | Mean duration (median) | Deer | Marked | |
|---|---|---|---|---|---|---|---|---|---|---|
East Limestone (2011) No-risk | 4 (15) | 14 | 8 | 7 | 7.7 (3.5) | 6 | 1.4 (0.3) | 3 | 3 | |
Kunga (2011) No-risk | 4 (12) | 91 | 56 | 42 | 5.2 (2.9) | 35 | 4.0 (1.4) | 28 | 7 | |
Kunga on Reef (2014) | 5 (16) | 31 | 6 | 6 | 4.7 (4.5) | 25 | 0.9 (0.6) | 5 | 5 | |
Native Reef (2011) Post-culls | 8 (35) | 9 | 6 | 0 | 1.2 (0.4) | 25 | 2.25 (0.3) | 18 | 4 | |
Native Reef (2014) Post-culls | 5 (16) | 35 | 13 | 4 | ||||||
Response to traps
On Reef Island (Post-culls) the capture rate was significantly lower than on islands without hunting (p ≤ 0.05) (Table 3). On Reef only 2/8 deer were captured more than once, compared to 35/51 for the No-risk islands, with a majority of individuals captured more than three times (Fig. 5).
Table 3 - Summary of box-trap capture data of animals on their native island for all sessions. Captures + recaptures = total number of captures including multiple recaptures of individuals within a session. Adjusted for outliers = for deer recaptured over 5 times within a capture session (ranged from 6 to 23 times) we capped their recapture scores in the session at 5 to limit variance inequality. This adjustment was conservative as it reduced the contrast between the no-risk and risk categories. Significance in differences in adjusted rates: East Limestone versus Reef: t = 3.31, df = 3.19, p = 0.04; Kunga versus Reef: t = 2.84, df = 3.71, p = 0.05; East Limestone versus Kunga: t = 1.06, df = 3.50, p = 0.17.
Captures+recaptures | Adjusted captures | |||||
|---|---|---|---|---|---|---|
Island | Years | Effort boxes*days | Nb. Deer captured | All | Adjusted for outliers | Trapping rates deer/box*day/year |
East Limestone | 2011- 2013 | 494 | 27 | 207 | 140 | 0.30±0.13 |
Kunga | 2011- 2013 | 591 | 23 | 150 | 102 | 0.16±0.07 |
Reef post culls | 2011- 2013 | 664 | 8 | 23 | 18 | 0.04±0.02 |
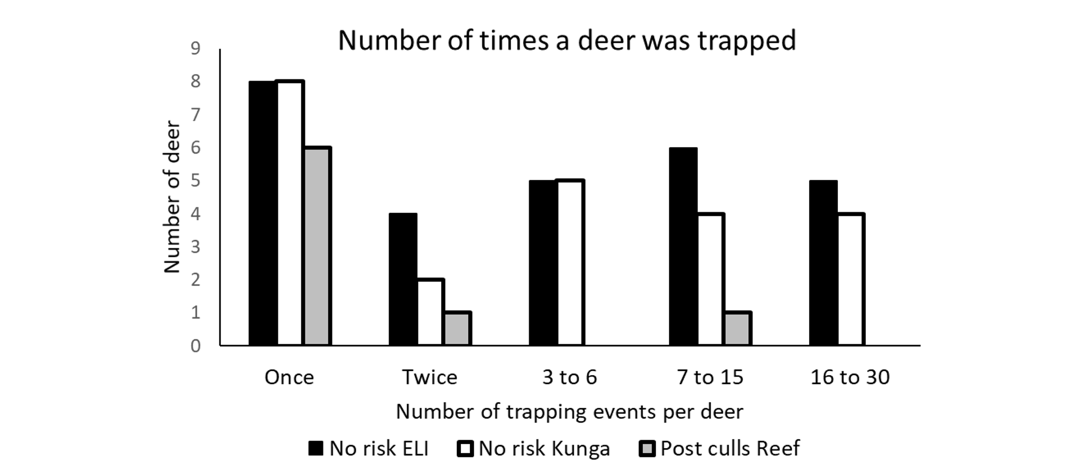
Figure 5 - Summary of the capture and recapture histories for the deer native to the three sites and two categories of hunting histories. ELI = East Limestone Island.
Diel activity pattern
Between 2011 and 2014 we recorded 762 (Table 4) picture sequences of deer on automatic cameras. We used the time recorded on the first picture in a sequence to assign the sequence to day time or night time. These pictures were taken in spring and early summer, hence a period of the year of long days and short nights. Deer were more active by day than by night (Table 4). There was no significant difference in daily activity patterns between East Limestone and Kunga islands (Chi-squared = 0.29, df = 1, p = 0.59) but deer native to Reef post-culls were more nocturnal than either (Chi-squared = 10.38, df = 1, p = 0.005). There was no significant difference in diel activity between marked and unmarked deer either on Reef or Kunga (Chi-squared = 0.05, df = 1, p = 0.81).
Table 4 - Distribution of deer observations by automatic cameras across sites and their assignment to day or night with day defined as the period between start of civil-twilight in the morning and end of civil-twilight in the evening. Night was defined as the period between civil twilight end and civil twilight start the following day.
Island/hunting history | N | day | night | % at night |
|---|---|---|---|---|
No-risk East Limestone | 289 | 256 | 33 | 11% |
No-risk Kunga | 358 | 311 | 47 | 13% |
Post-culls Reef | 115 | 88 | 27 | 24% |

Observer monitoring an unruffled wild deer foraging on remote Kunga Island (No-risk).
Shoreline use by deer varied with risk and forest understory vegetation
The following diagnostic and tests confirmed that our Markov Chain Monte Carlo (MCMC) chains converged in the MixSiar model. The Gelman-Rubin Diagnostic run after the MixSiar model indicated that only 2 of the 140 variables exceeded marginally the threshold value of 1.01. The Geweke diagnostic, a standard z-score to assess if less than 5% of the 140 variables were outside the +/-1.96 values in each chain indicated 3%, 5% and 1% for chains 1, 2 and 3 respectively.
On the isospace plot (Fig. 6) the isotopic signature in deer bone samples were distributed in relation to the proportion of seaweeds in deer diet relative to terrestrial plants. Samples from the no-risk islands had a higher proportion of seaweeds in deer diet than samples from sites with a risk history (Fig. 6b). The samples from Murchison and Faraday showed the highest mean proportion of seaweeds, those from Graham and from the BC Central Coast, where hunting and/or predators occured, the lowest (Fig. 6b). Among the terrestrial plant groups the cluster which included deciduous trees and shrubs, ferns, forbs and grasses, made up the highest proportion in deer diets. Conifers came next and salal last.
The MixSIAR analysis quantifies the proportion of the four plant categories in deer diets in the samples (Figs. 6 and 7). Bayesian 95% credible intervals (Fig. 8) allow us to estimate the significance of differences in the proportion of seaweeds between the samples. Credible intervals from the samples without predation (Murchison, Faraday, East Limestone, Kunga and Reef before the culls) do not overlap with those of the two sites with hunting and/or predators (Graham and BC Central Coast). The credible interval of the Post-culls Reef sample was intermediate.
The proportion of seaweed in deer diets reached 31% on Murchison and Faraday, and 19.2% on sites in Laskeek Bay (East Limestone, Kunga and Reef prior to the culls). It dropped to 8 and 9.6% respectively in sites with hunting (Graham) or with predators and hunting (BC Central Coast). On Reef after the culls the proportion of seaweed in diet (13.5%) was intermediate between the no-risk value and the values for the hunting or predators+hunting samples (Figs. 7 and 8).
The proportion of the terrestrial plants in deer diets, which included deciduous trees and shrubs, ferns, forbs and grasses varied from 67% on Murchison and Faraday to 88% on Graham and on the coastal mainland. Conifers and evergreen shrubs stand out as a minor component of diet. It was always lower than 8% for conifers, despite their prevalence in the vegetation profiles of some of our study sites (Fig. 3). The proportion of salal (evergreen shrub) never exceeded 4% in the diets.

Deer feeding in the intertidal on a risk-free island on Haida Gwaii
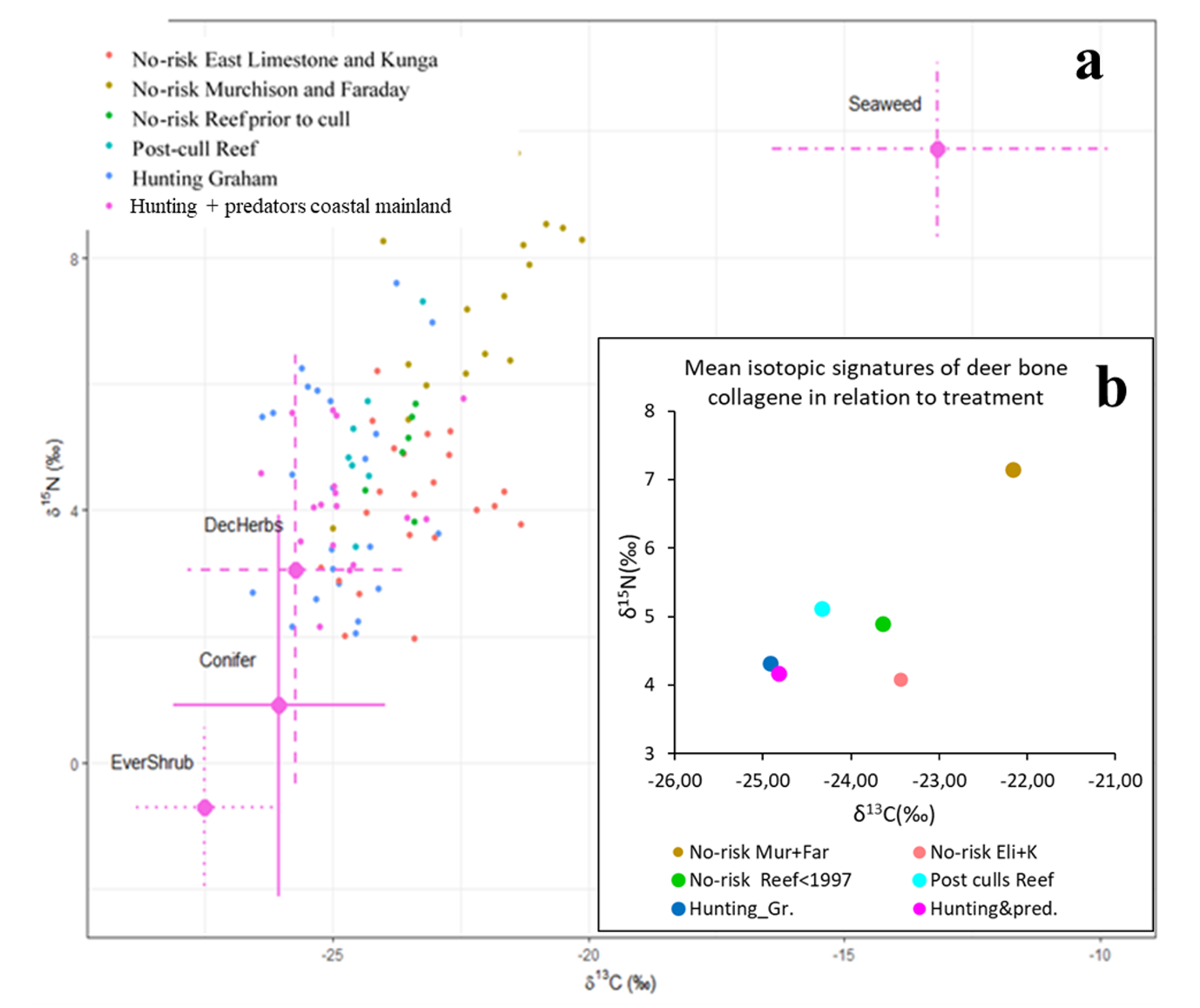
Figure 6 - Isospace plots of deer bone collagene and plants, Mixture data by risk category and islands. No-risk Murchison and Faraday = No-risk samples from Murchison (N = 10 bone samples) and Faraday (N = 6); No-risk samples from East Limestone (N = 10) and Kunga (N = 12); No-risk samples from Reef Island prior to the culls (N = 7); Post-culls Reef = samples from Reef Island after the culls (N = 6); Hunting Graham = samples from Graham Island (N = 23); Predators + hunting = samples from the BC Central Coast (N = 17). Source data are by risk/island categories and have been adjusted by discrimination means and SDs. Terrestrial plants: EverShrub = evergreen shrub (Salal), Conifer = conifers, DecHerbs = deciduous trees and shrubs, forbs, ferns and grasses; Seaweed = Seaweeds (40 species). Error bars indicate 1 SD, the combined source+discrimination SD calculated under assumption of independence as: \(\sqrt{\sigma^{2}source + \sigma^{2}discr}\)
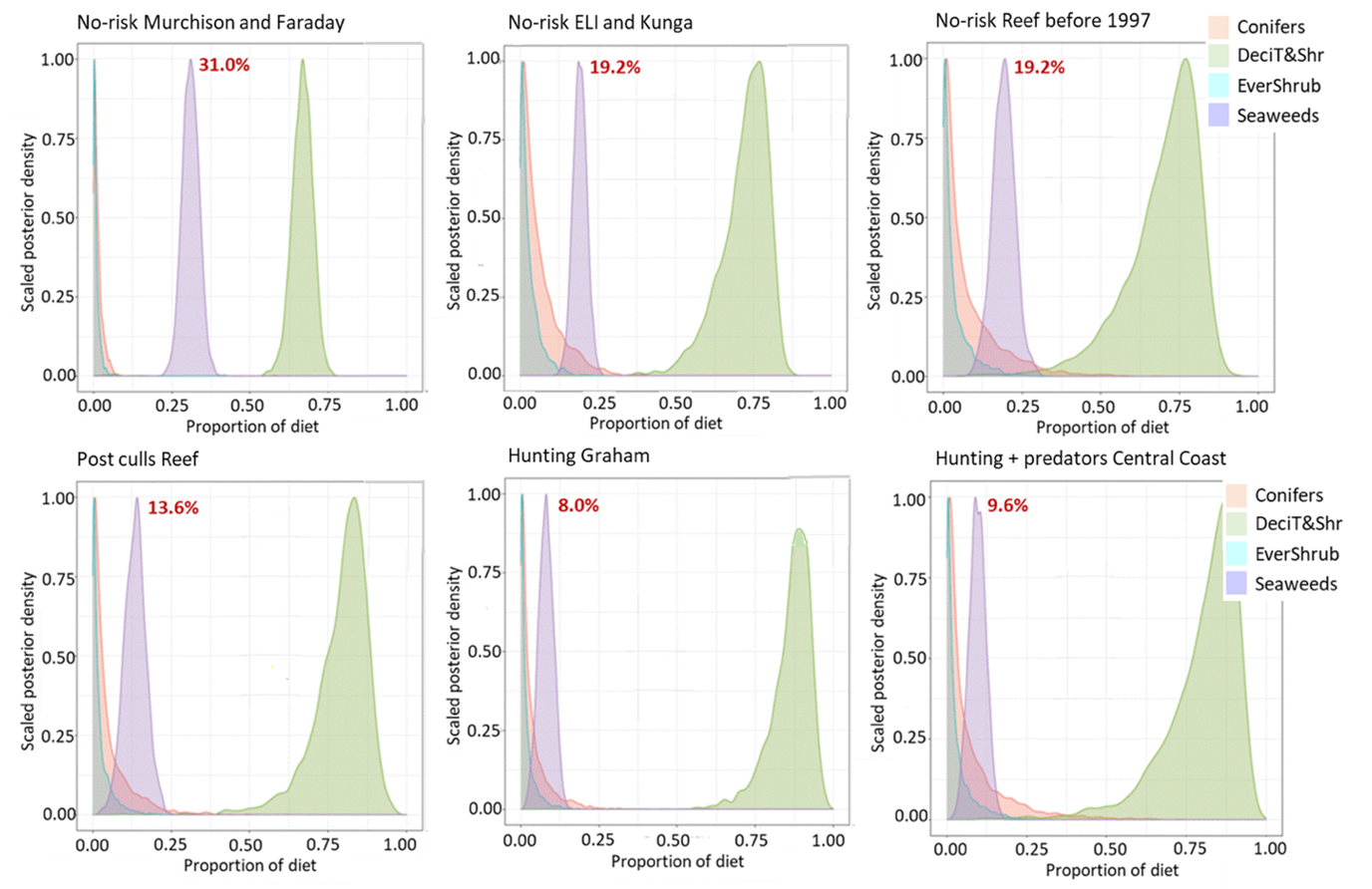
Figure 7 - MixSiar posterior plots of overall deer diet in relation to predation risk. N = number of bone samples per site. No-risk history, four sites: Murchison (N = 10), Faraday (N = 6), East Limestone (ELI) (N = 10), Kunga (N = 12) and Reef before the culls (N = 7)) (see map in Fig. 1); Post-culls Reef = samples from Reef Islands collected after the culls (2008-2014) (N = 6); Graham Island coastal area = yearly relatively low intensity hunting without marked effect on high deer density (Engelstoft, 2001) (N = 23); BC Central Coast, hunting and predators (N = 17). All profiles are at the same scale. Murchison and Faraday are two islands with vegetation poor understories [see text and Martin, Gaston & Hitier (1995)]. The peaks of the shaded areas represent the median estimate of the proportion of the different sources in deer diet. Pink shading = conifers, 4 species [Sitka spruce, western hemlock, western redcedar, and yeellow cypress (Chamaecyparis nootkatensis)], 13 samples; light blue shading = evergreen shrubs, 1 species (salal), 4 samples; green shading = deciduous trees and shrubs, forbs, ferns and grasses (27 species, 86 samples); purple shading = seaweeds (40 species, 237 samples). Red figures indicate median percent value for seaweeds.
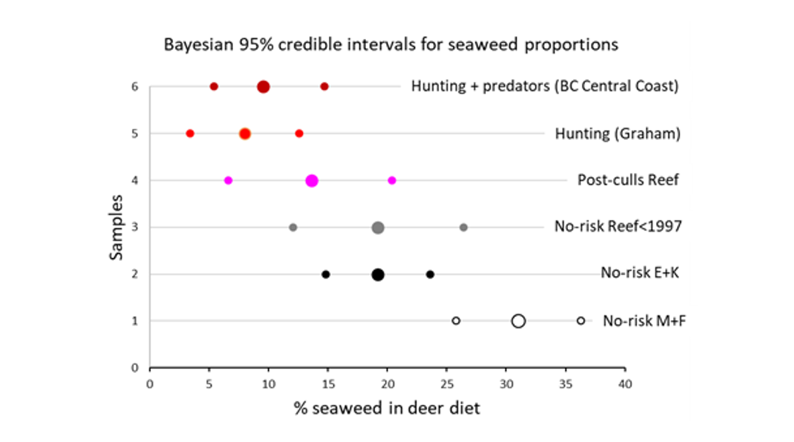
Figure 8 - 95% Bayesian credible intervals of the 50% quantile (large central dots = median proportion of seaweeds in diet) as provided by the 2.5% and 97.5% quantiles (small dots). M+F= Muchison + Faraday islands; E+K = East Limestone + Kunga islands.
Discussion
How did understory vegetation cover and composition change with predation risk?
Our results illustrated the well-established effect that large herbivores can have on forest vegetation cover (Côté et al. 2004; Martin et al. 2010, 2011; Cardinal et al. 2012; Chollet et al. 2013, 2021a; Waller & Reo 2018). They also illustrated the relationship between the level of impact on the vegetation and the risk history a deer population has been exposed to (Callan et al., 2013; Waller & Reo, 2018). The severe culls on one of our sites (Reef Island) resulted, over a decade, in an upsurge of understory vegetation and regenerating conifers in an initially heavily-browsed open understory (Figs. 2 and 3), combined with an increase in diversity among plots [(Fig. 2 and Chollet et al. (2016)]. However, there are intrinsic differences between this recovering understory vegetation and the understory vegetation we sampled on Yeo Island where deer have always been subject to their natural predators in addition to hunters. On Reef, the high proportion of regenerating conifers in the understory following the culls resulted from the release of heavily browsed dwarfed spruce and hemlock (Chollet et al. 2016), while the high cover of grasses resulted from a rapid colonisation of bare ground (Chollet et al. 2016). By contrast, the vegetation profile we observed in the presence of large carnivores and hunting on Yeo was remarkably similar to the understory vegetation profile we observed on three islands in Laskeek Bay never colonized by deer [see details in (Stockton et al. 2005, Martin et al. 2010)]. Their understories lack the significant cover of conifer regeneration we observed on Reef after the culls and are dominated by shrubs and forbs (Fig. 3).
On Graham Island, where hunting is of low intensity and restricted to sites most accessible to the small local population of hunters (Martin & Baltzinger 2002), the diversity of cover among plots (spread of plots, Fig. 2) and the amount of shrub cover (Fig. 3) was higher than on No-risk islands but understory vegetation overall was still closer to the one observed on No-risk islands [for more details on the understory vegetation on Graham see (Engelstoft, 2001; Engelstoft et al., 2008; Chollet, Baltzinger, et al., 2021)]. This variation suggests an interplay among the presence of risk, its intensity, its nature (natural predators present or not) and its duration in the interaction between deer and their habitat.
Does absence of risk favor boldness and deer impact on the vegetation?
Our comparison between populations that had not been exposed to any risk (East Limestone and Kunga), with a population on Reef subjected, over a decade prior to the study, to a severe population reduction through culling, revealed clear behavioural contrasts. In absence of risk deer were easy to trap and unwarry of people. They also willingly consumed foreign food at bait stations, and foreign food when associated with an unfamiliar object (trap). The significantly shorter flight initiation distances, and much shorter distances travelled, observed in non-hunted populations were retained by the Kunga deer translocated to Reef, even though they showed slightly longer flight initiation distances in their novel environment. This was in stark contrast with the wary behaviour of the native deer on Reef after the culls, even more so as the flight and movement distances we recorded for the Reef deer were conservative: they often fled before being seen (heard running) and others were still running when they disappeared from view. As a result, actual values on Reef may have been closer to values reported for black-tailed deer populations on the mainland (FID ~ 60 to 70 m and Dtravel ~ 70 to 120 m), where deer can be subject to hunting and carnivores (Stankowich & Coss, 2006, 2007; Stankowich, 2008). Our field notes also indicated that native deer on Reef ran or trotted away, whereas deer from Kunga, even after translocation, and East Limestone (No-risk islands) usually performed a slow walk to a nearby location where they resumed their activities. On East Limestone, FID and distances travelled were significantly shorter than on Kunga, possibly reflecting some habituation to human presence (Rodriguez-Prieto et al., 2009; Schuttler et al., 2017) on East Limestone Island, where people are present daily for two to three months each summer for ecological monitoring.
Despite minor differences in flight distances or time spent at bait, marked and unmarked deer of the same population had comparable flight initiation behaviours. Marked and unmarked deer from Reef born after the culls were predominantly shy, as their poor trapping rate illustrated, even when we take into account that deer densities on Reef at the time of study were still at only 50% of their pre-cull density (originally ~30 deer/km², i.e. > 80 deer present; Chollet 2012). Only two (25%) of the eight deer captured on Reef were ever re-captured, and only one was recaptured repeatedly, mostly as a yearling and a juvenile, before becoming trap shy in the following years while still captured on cameras. This contrasts with deer unexposed to risk on East Limestone and Kunga. A significant proportion of the local deer were trapped, and over half of them re-trapped three times or more over the years (Fig. 5). The presence on Kunga and Reef of a fraction of animals never trapped, but captured on automatic cameras, indicated that within each population there was variation in trap-shyness among individuals (Sih et al., 2012). Overall, observed differences in behaviour between deer native to East Limestone and Kunga and those native to Reef after the culls, suggest that culling shifted the dominant behaviour profiles of Reef Island deer towards the shy-end of a boldness-shyness continuum.
Food limitation and nutritional status have been proposed as mechanisms explaining individual attenuation in antipredator behaviours such as flight response (Stankowich, 2008; Gaynor et al., 2019). If so, we expect these behaviours to be reversed once food supply improves. Could a better food supply in the lusher understory on Reef after the culls explain the higher values of FID and Dtravel for deer native to Reef, and would the apparent paucity of resources in the heavily browsed understories of East Limestone or Kunga explain the apparent lack of fear in their deer? This seems unlikely, given that we did not observe any significant change in the behaviour of the deer translocated from Kunga to Reef (Fig. 4). Our observations rather suggest a more indirect link beweeen behaviour and resources, a link mediated by risk. Following the reasoning of Kirchhoff & Person (2008) we assumed that in absence of risk, intensive browsing would, over time, decrease the per capita availability of forage as well as the average quality of the diet. A reduction in anti-predator behaviour could be the necessary condition to allow increased foraging time in response to this progressive erosion of standing plant biomass and forage quality in an increasingly browsed understory. In these understories, resources, although sufficient to sustain a dense deer population (Stockton et al., 2005; Le Saout, Chollet, et al., 2014), are increasingly found in small increments of emerging vegetation or as subsidies fallen from the canopy (Le Saout, Chollet, et al., 2014). Indeed, on No-risk islands less then 5% of the resources needed by these dense deer populations were provided by the rare standing biomass. The bulk was provided equally by the cryptic flux of growing vegetation and by canopy foliage fall, supplemented by seaweeds from the shorelines (Le Saout, Chollet, et al., 2014).
Our results from the stable isotope analysis are another illustration of the focus deer have to put on cryptic vegetation elements in the understory. On one hand conifers, the dominant element in the understory of No-risk sites (Fig. 3), make for only a tiny proportion of their diet (Fig. 7), whereas deciduous vegetation, a known preference in black-tailed deer diet (Taylor, 1956; Pojar, 1999), make up the bulk of their diet (Fig. 7), its negligible presence as standing biomass (Fig. 3) compensated by short lived emerging vegetation (Le Saout, Chollet, et al., 2014). This need to feed on spatially dispersed food, occurring as small items, could be the ecological context forcing the emergence of populations expressing less anti-predator behaviours (Réale et al., 2010; Sih et al., 2012), or with less costly adjustments of spatial distribution in response to perceived distribution of risk (Williams et al., 2008; Gaynor et al., 2019). Such behavioural shifts could, in turn, help deer intensify their foraging pressure and, over time, aggravate the loss in understory standing vegetation, as was actually shown on these islands between 1989 and 2009 (Chollet et al., 2015). This could further increase the shift towards bolder behaviours. The overall predominance of diurnal activity in East Limestone and Kunga deer compared to populations exposed to hunting (Bonnot et al., 2016) can also be interpreted as a change in favour of increased foraging time. This more diurnal behaviour has been shown to contrast with the more nocturnal behaviour commonly observed in hunted populations (Altendorf et al., 2001; Bonnot et al., 2016, 2020; Palmer et al., 2022). However, anti-predator behaviours less costly in foraging time such as routine vigilance would be retained and not counter-selected as was actually observed for the populations of East Limestone and Kunga where routine vigilance persisted in an otherwise unwary population. (Chamaillé-Jammes et al., 2014; Le Saout et al., 2015).
On Reef, after the first few days of culling, every animal remaining was much shyer than the animals observed pre-cull (AJG and T. Husband pers.com), showing that at least some animals in the pre-cull population, previously never hunted and living in a heavily browsed habitat for more than 50 years (Martin et al. 1995; Vila et al. 2004a; Chollet et al. 2016), had retained the ability to respond to a new threat [see also Chamaillé-Jammes et al. (2014)]. A study on the effects of low intensity and simulated hunting (Le Saout, Padié, et al., 2014) on the behaviour of deer on Kunga also documented the presence within the local deer population of individual personalities differing in their ability to respond to a threat. Some animals were easily seen and trapped, and showed little or no response to simulated hunting or to being trapped, whereas some animals were only observed through automatic cameras, were never trapped and shifted their habitat use away from the area exposed to simulated hunting.
On Reef, at the time of this study, the animals born several years later, offspring of those that survived the culls, were much shyer than those seen on Reef prior to the cull (AJG pers. obs.). These animals also had a tendency to be more active at night than deer on Kunga and East Limestone never exposed to hunting or to culls. The survivors on Reef may have included animals that modified their behaviour more rapidly and more radically than those that were killed. If some or all of that variation in behaviour was under genetic control (Réale et al., 2007), subsequent generations will have been shyer than the pre-cull population as a result of differential survival of shyer personalities during the culls. The consequent persistence of increases in FID, Dtravel, novelty aversion and nocturnal foraging would have been inherited or adopted by young reared subsequently, so that the behaviour of the current population resembles that of populations subject to natural predation and human hunting. Rapid differential selection of heritable antipredator behaviours in response to novel predation were documented recently (Moseby et al., 2023). Conversely, there is little sign that the deer trapped and translocated from Kunga Island and hence never exposed to predation threat, adopted any of the modified behaviour of the local post-cull deer despite the better foraging resources.
Did risk affect the use of habitats?
The lower proportion of seaweeds in diets at sites with predators and/or hunters seems to validate our prediction that deer would make less use of exposed shorelines where they face predation from hunters and/or wolves. But, as the amount of standing vegetation in the understories tends to be much lower at sites without hunting (Fig. 3) it could be the lack of forage in the underestories of No-risk islands that encouraged deer to forage in the intertidal. On Murchison and Faraday, where seaweed is especially prominent in the diet, the very open understories below dense secondary canopy, supported very little ligneous and herbaceous vegetation cover in the 0 to 1.5 m layer (< 4%, Martin et al. 1995).
However, the two situations with the lowest and similar proportion of seaweed isotopes in jawbone collagen, Graham and Yeo Islands, differed in their cover of standing understory vegetation. On Yeo, the permanent presence of predators and hunting was associated with a lush understory. On Graham, the long history of moderate hunting, restricted to areas most accessible to a small number of hunters (Martin & Baltzinger 2002), had limited impact on deer density (Engelstoft, 2001; Engelstoft et al., 2008), resulting in understory cover closer to that at No-risk sites than to that at sites on the BC Central Coast (Figs. 2 and 8) (Pojar, 1999; Chollet, Baltzinger, et al., 2021). This partial decoupling of understory vegetation cover and proportion of seaweeds in deer diet remains suggestive that risk history on a site can negatively affect deer propensity to feed on shorelines. This would be consistent with the documented high use of shorelines by foraging wolves and its negative consequence on their use by deer (Klein, 1995). It would also be consistent with the documented increased reluctance in deer to use more exposed habitats or localities when risk is higher (Williams et al., 2008; Bonnot et al., 2017).
What lessons for ecology, population behavioural change and conservation?
Our research contributes to the limited number of studies that connect animal behaviour with ecological dynamics, species interactions, population changes and ecosystems [see (Sih et al., 2004, 2012; Réale et al., 2007; Chitwood et al., 2022)]. We show that the introduction of a new threat, such as hunting on Reef Island, led to a sudden change in behaviour. This change left the survivors exhibiting behaviours that were significantly different from those of animals on islands without hunting.
Our findings support the hypothesis that shifts in the “ landscape of fear” (Gaynor et al., 2019; Palmer et al., 2022) can change the behavioural traits of a population. These changes may occur in responce to intense selective pressures on particular behavioural traits caused by immediate risk (e.g. as in the case of severe culling), or, over longer periods of time, from the progressive reproductive advantage of behavioural profiles better adapted to a changing environment (e.g. a progressive increase in boldness driven by a progressive depletion of resources in absence of acute predation risk). Vourc’h et al. (2001, 2002b) documented similar shifts in population profiles in redcedars exposed or not to browsing by deer on Haida Gwaii. Trees with low levels of chemical defenses dominated in populations not exposed to browsing, while individuals with high levels of defenses dominated populations exposed to deer. These defence levels were under genetic control (Vourc’h, Russell, et al., 2002; Vourc’h, Vila, et al., 2002).
While our interpretation of the non-wary behaviours we observed on islands without predation or hunting, or of the rapid shift towards shyness after the culls, remains speculative, it highlights the possibility for behaviour selection. The speed of behavioural change will be influenced by the intensity of the selective pressure (Blumstein, 2002; Jolly et al., 2018; Moseby et al., 2023). It can be rapid if severe culling continues over generations but may be slower in the case of the relaxation of antipredator behaviours after the colonisation of a predator free environment (Blumstein, 2002). The evolution of predator naivety in island organisms provides many classic examples of both gradual and rapid evolutionary changes in behaviour (Darwin, 1840; Lack, 1968; Blumstein, 2002), but see Blumstein & Daniel (2005).
Our results have implications for conservation. They can improve understanding of the problems posed by the adjustment of focal species towards a novel threat, thus improving the success of species restoration programs where naïve captive-bred individuals have to adjust to predators on release (Moseby et al., 2015, 2016, 2023). They also provide insights into the management of overabundant populations through hunting. In such instances, a better grasp of behavioural responses to management actions can help design strategies that take into account the ability of species to adjust their spatial distribution and diel rhythms to the threat posed by management measures (Williams et al., 2008; Potratz et al., 2024). A better understanding of the ramifications of consequences triggered in prey by the return of their predators will also help better address the challenges posed by high deer populations in parts of Europe and North America to human activities, such as farming, livestock husbandry, forestry (Kuijper et al., 2013, 2016; Raynor, 2017; Martin et al., 2020) or road safety (Gilbert et al., 2017; Raynor et al., 2021; Bell et al., 2024). Finally, by improving our understanding of mechanisms that lead to evolutionary shifts, our results help to better predict the full impact of neglecting the integrity of species assemblages. Hence, our evidence encourages the conservation and restoration of fully functioning ecosystems.
Ethical note
All research was conducted under the appropriate animal care permits from the Wildlife Act of the Ministry of Natural Resources Operation of British Columbia (No. NA11-68421, approved by Parks Canada Animal Care Task Force research permit No. 9059), and under the Archipelago Management Board of Gwaii Haanas National Park Reserve and Haida Heritage Site research permit No. GWA-2011-8373. All except 3 deer captures were done using box traps specially designed for this project (the exceptions were one individual captured by a clover trap and two by a netgun). We made over 400 deer captures or recaptures in the course of the project. Traps were radio-monitored remotely and we dealt with captured animal immediately after capture notification. The animals captured at night were treated the following morning. We recorded no injury to deer, nor mortality associated to capture. The project permits included the experimental culls on one island and the translocation of a small number of deer from one island to another. Culls were conducted by local hunters under clear ethical and practical rules. Carcasses were processed and the meat distributed to the local communities.
Acknowledgements
This is a publication of the Research Group on Introduced Species (RGIS, https://rgis.cefe.cnrs.fr). Preprint version 6 of this article has been peer-reviewed and recommended by Peer Community In Ecology (https://doi.org/10.24072/pci.ecology.100643; Chaverri, 2024). On top of funding (see below) we received critical logistic support from the Laskeek Bay Conservation Society, Environment and Climate Change Canada, the Gwaii Haanas National Park Reserve, National Marine Conservation Area Reserve and Haida Heritage Site, the Coast Sustainability Trust Matching Fund (CST-QCI-HG- 061– RGIS Infrastructure Upgrade), and the “Understanding Canada” program from the Canadian government. We are indebted to Todd Golumbia, Carita Bergman and Peter Dyment and colleagues from Gwaii Haanas for support in the field. Tibo Verchère, Ainsley Brown, Barbara and Keith Rowsell, Greg Martin, Jacques Morin, Erin Harris, Terry and Ron Husband, Charlotte Tarver, Mike Gillingham, Malcolm Hyatt, Georges Janeau, Jean-Luc Rames, Denis Picot, Nicolas Morellet, Bruno Cargnelutti, Nicolas Cebe, Helen Schwantje, Tom Smith and Keith Tipper provided critical help in the field and beyond. Chris Darimont, Eva Poilvé, Hélène Verheyden, and Maurah Van Impe assisted with the treatment of samples in the stable isotope analysis. The Haida Watchmen program provided precious help through accommodation on Tanu Island. We are deeply thankful towards the heredititary chiefs of T’aanuu Llnagaay (Tanu) and Ḵ'uuna Llnagaay (Skedans) for welcoming us on their land. Over the years, many people from all walks of life of the local communities provided invaluable support. We are deeply thankful to all. We also benefited from very insightful suggestions by two anonymous reviewers and by the recommender Gloriana Chaverri.
Data and scripts availability
Funding
Funding for this research included grants from the French Ministry of Foreign Affairs, from the French National Agency for Research (#2010-BLAN-1718, BAMBI), the International Research Group (GDRI) Dynamics of Biodiversity and Life-History traits from CNRS, the South Moresby Forest Replacement Account (SMFRA) and Gwaii Forestry Charitable Trust (GF), and from National Geographic # 9266-13.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article. Jean-Louis Martin and Simon Chamaillé-Jammes are listed as recommenders.