
 CC-BY 4.0
CC-BY 4.0
Introduction
Coevolution between plants and their pollinators is believed to be a major driver of floral traits diversification in animal-pollinated angiosperms (Darwin, 1862; Fenster et al., 2004; Harder & Johnson, 2009). The increasing number of experimental studies evidencing the occurrence of pollinator-mediated selection on floral traits (Caruso et al., 2019), along with studies linking pollination mode to floral diversification in a phylogenetic framework strongly support this driving role of plant-pollinator interactions in shaping floral evolution, particularly in specialized plant species (e.g Graham & Barrett, 2004; Whittall & Hodges, 2007). It should be noted, however, that only a minority of plant species rely on such highly specialized pollination systems (for instance plants that are engaged in brood-site mutualisms, e.g. Pellmyr, 1992, or plants that rely on sexual mimicry to attract their pollinators, e.g. Peakall et al., 2010). Indeed, studies documenting pollinator assemblages often show that plant species are visited by multiple potential pollinator taxa (Kato, 2000; Zhang, 2017), and the relative role of each visitor as a selective agent is generally not clear. This role is likely to depend on several parameters of the plant-pollinator interaction, in particular (i) the relative abundance of that pollinator in the local community, (ii) its visitation rate and pollination efficiency (defined as the amount of pollen transported and deposited on the stigmas of flowers visited later in the sequence; Wu et al., 2018; Caruso et al., 2019), as well as (iii) the effect that floral traits have on its visitation rate and pollination efficiency. Indeed, different pollinator species can differ in the floral traits that are used as attractive signals (Schemske & Bradshaw, 1999), or differ in their preference for a given trait value (Hoballah et al., 2007; Gómez et al., 2008; Gong & Huang, 2009).
In this regard, plants with mixed pollination systems including both nocturnal and diurnal pollinators constitute interesting study objects, because both groups of floral visitors are not necessarily attracted by the same signals. In particular, it is generally expected that visual signals should be crucial to attract diurnal pollinators, while fragrance should be more important for nocturnal pollinators (Fenster et al., 2004). For instance, nocturnal hawkmoths have been shown to be mainly attracted by olfactory cues, whereas diurnal hawkmoths seem to be preferentially attracted by visual cues (Balkenius et al., 2006). In plants with mixed pollination systems, overall selection on floral traits will thus be the product of different selection pressures mediated by diurnal versus nocturnal pollinators, reflecting their preferences for certain floral traits and their pollination efficiency.
Exclusion experiments, where plants are exposed to only one type of pollinator, enable us to investigate how exposure to different assemblages can impact various aspects of plant reproduction. Pollination by nocturnal versus diurnal assemblages can affect plant reproductive success, with the direction of the effect depending on the plant species (Giménez-Benavides et al., 2007; Sletvold et al., 2012; Amorim et al., 2013; Stone & Olson, 2018; Vanderplanck et al., 2020; Jaca et al., 2020). Foraging behaviour and pollination efficiency can indeed vary greatly between nocturnal and diurnal pollinators. A recent study of a generalist plant species (Rubus futicosus) showed that despite lower visitation rates, pollen deposition rates were higher with nocturnal pollinators than diurnal pollinators (Anderson et al., 2023). This suggests that the identity of pollinators could affect, in addition to reproductive success, the number of reproductive partners that a plant can acquire (i.e. the mating success). Moreover, pollen dispersal distance has been estimated to vary according to the type of pollinators, with nocturnal pollinators dispersing pollen further in some systems (Young, 2002; Barthelmess et al., 2006), which could also affect both mating and reproductive success.
Pollination systems have been widely studied in the Silene genus, and species have traditionally been described as either nocturnal, diurnal or mixed based on their floral traits and on the circadian rhythm at which their flowers are open (Greuter, 1995; Jürgens et al., 1996; Prieto-Benítez et al., 2015). In that genus, the supposed pollination system can vary even between closely related species. For instance, while in S. latifolia nocturnal pollinators - mainly moths - are indeed more efficient, with relatively higher seed-set during nocturnal pollination compared to diurnal pollination (Young, 2002; Scopece et al., 2018), the pollination system remains more ambiguous for S. dioica, our study species (Jürgens et al., 1996; Waelti et al., 2009; Prieto-Benítez et al., 2015). Indeed, S. dioica (i) is visited by diurnal pollinators but also nocturnal moths (Jürgens et al., 1996), (ii) can be predated by larvae of Hadena species, a moth genera involved in a nursery pollination interaction with many Caryophyllaceae species (Kephart et al., 2006; Prieto-Benítez et al., 2017) including the sister species S. latifolia (Dufaÿ & Anstett, 2003; Bopp & Gottsberger, 2004), and (iii) emits comparable amounts of scent during night and day (Waelti et al., 2008). Despite these observations, S. dioica is generally described as a diurnal species in the literature (Jürgens et al., 2002; Jürgens, 2004), primarily because, unlike its sister species S. latifolia, its pink flowers remain open throughout the day.
Investigating the exact impact of floral scent on plant reproductive success via its effects on pollinator attraction is challenging, since floral scents are usually complex blends of many different volatile organic compounds (VOCs), and both identity and relative proportions can constitute an attractive signal for specific pollinators (Bruce et al., 2005; Raguso, 2008; Proffit et al., 2020). In this context, phenotypic manipulation is a useful tool to investigate the effects of isolated VOCs on pollinator attraction (Campbell, 2009; Landolt et al., 2013). For instance, the role of scent in pollinator attraction can be studied by artificially increasing the emission of one molecule that is known to be produced by the study plant and has documented effects on pollinator attraction. One VOC in particular, phenylacetylaldehyde (PAA, Benzenoid), is known to be relatively widespread among angiosperms (Schiestl, 2010) and has been described to be of particular importance in moth attraction (Cantelo & Jacobson, 1979; Heath et al., 1992; Tóth et al., 2010). Several lines of evidence suggest that PAA can also be implicated in diurnal pollinator attraction: (i) butterfly species show a strong antennal responses to increased quantities of PAA (Andersson & Dobson, 2003), (ii) bumblebees have been shown to be able to develop a preference for this VOC when associated with reward (Knauer & Schiestl, 2015) and (iii) an experimental evolution study demonstrated that Brassica rapa populations showed an increase in PAA emission across generations when exposed to bumblebees (Gervasi & Schiestl, 2017). Although low compared to some other Silene species, in particular those described as being night-pollinated (Jürgens, 2004; Page et al., 2014), PAA can be found in moderate amounts in floral scent of S. dioica and is released in similar amounts during the day and the night (Waelti et al., 2008). This raises the question of how the emission of an attractive VOC impacts diurnal versus nocturnal pollinator attraction and plant reproductive success. Finally, PAA emission could interact with selection on the other attractive floral traits, if PAA enhances diurnal or nocturnal pollinator attraction. Indeed, this could weaken selection on other traits because scent emission would be a more important feature to pollinators (thus effectively removing the selective advantage of other attractive floral traits). On the contrary, PAA emission could strengthen selection on other traits by enhancing visits by specific pollinators, which also use floral traits as visual cues when visiting the plants.
In this study, we explore the effect of diurnal versus nocturnal pollination on male and female mating and reproductive success in dioecious S. dioica, as well as the effect of an artificial increase in PAA emission on selection on floral traits, in a fully-crossed design. We ask the following questions: (i) Are individuals more limited in their mating and/or reproductive success at night because of limited pollinator visits and/or predation by Hadena bicruris? (ii) Does an increase in PAA emission impact reproductive success (i.e. through better pollination and/or more predation), and does this effect vary between diurnal versus nocturnal pollination? (iii) Are the same attractive traits under selection when plants are exposed to diurnal versus nocturnal pollination? (iv) Does an increase in PAA emission affect selection gradients on floral traits? (v) How is pollen dispersal distance affected by diurnal versus nocturnal pollinators foraging behavior?
Material and methods
Study system and plant material
Silene dioica (L.) Clairv. is a dioecious short-lived perennial species of the Caryophyllaceae family. It is widely distributed throughout most of northern and central Europe (Baker, 1947; Jalas & Suhominen, 1986). Sexual dimorphism in floral traits is prevalent in this species, with males exhibiting larger flower sizes, greater flower numbers, and longer flowering durations, whereas females produce more nectar per flower (Kay et al., 1984; Hemborg, 1998; Moquet et al., 2020; Barbot et al., 2023). It has a generalist pollination system and is thought to be mainly pollinated during the day (Jürgens et al., 1996; Kephart et al., 2006), with Bombus species and Syrphidae described as main pollinators (Baker, 1947; Westerbergh & Saura, 1994; Barbot et al., 2022), but Noctuidae species have also been shown to act as pollinators (Jürgens et al., 1996). Beyond their role as pollinators, nocturnal pollinators of the genus Hadena are also recognized as predators of Silene dioica (Prieto-Benítez et al., 2017). This interaction between Hadena moths and Silene dioica flowers occurs within a brood pollination system, where adult moths not only pollinate the flowers but also deposit their eggs inside them. The emerging larvae then feed on the reproductive tissues of the host plant. (Kephart et al., 2006). Flowers of Silene dioica emit scents dominated by benzenoids (including PAA) and monoterpenoids, both during day and night (Waelti et al., 2008).
Experimental population and common garden
The experiment spanned seven days, from July 6th to July 12th, 2019, and was set up in a common garden on the campus of Lille University in France (50°36'27.9’’N 3°08'36.3’’E), several kilometers away from the nearest wild populations of Silene dioica. The experimental population was created using the same collection of plants as in Barbot et al. (2022). All individuals were in separate 0.7-L pots filled with a standard soil mixture, and were repotted each year. In S. dioica, males flower for a long time (up to 2 months), while females have a shorter flowering period, generally around one month. Female flowering is characterized by slow flower production at the beginning and end, with a marked peak in the middle (Moquet et al., 2020; Barbot et al., 2023). For this experiment, we selected female plants that were at a similar stage in their flowering phenology, with 11.22 (± 9.53 SD) open flowers on average and many flower buds, indicating they were approaching their peak flowering stage.
Experimental treatments
We used a fully-crossed design to investigate the effect (i) of diurnal (D) versus nocturnal (N) pollination and (ii) of unmanipulated (C) versus increased (T) phenylacetylaldehyd (PAA) emission on mating and reproductive success, with 30 females and 30 males per combination of treatments (i.e. DC, DT, NC and NT). Individuals were kept in an insect-proof greenhouse until the beginning of the experiment. All plants were randomly assigned to one of the four groups before the experiment. The surface area of the experimental plot was of 31.35 m-2 (5.5 × 5.7 m) and plant density was 3.75 individuals.m-². In the experimental garden, for each pollination exposure treatment, individuals were spatially arranged in order to alternate (i) females and males and (ii) C and T treatments. In addition to the plants included in the experimental design, 16 additional females were randomly selected from the same collection and hand-pollinated (HP plants, see details of treatment below). HP females were placed in the same experimental garden, five meters away from the experimental plot.
Pollinator exclusion treatments
Each morning and evening, plants were moved between an insect-proof greenhouse and the experimental plot where they could be visited by the local pollinator fauna: (i) plants in the diurnal pollination group (D) were placed outside on average from 6:30 am to 9:30 pm and (ii) plants in the nocturnal pollination group (N) were placed outside on average from 9:30 pm to 6:30 am. Exposure timing reflected shifts in pollinator communities in the experimental garden (pers. obs.).
PAA manipulation
Preliminary experiments were conducted to design a protocol allowing PAA emission in the T group to be twice the total average emission of all VOCs described for S. dioica in the literature (110 ng.h-1 per flower, with 20 flowers on average per plant, thus 2200 ng. h-1 per plant, Waelti et al., 2008). In all plant pots (i.e. both C and T plants), we planted a wooden stick supporting a glass tube containing 4mL of paraffin oil. In plants from the T group, a 95% PAA solution was added (1:400 dilution in the paraffine). Tubes were then sealed and a 1µL micro-capillary tube was inserted in each glass tube in order to allow slow diffusion. Tubes were finally insulated with aluminum foil in order to minimize the differences in diffusion between day and night due to differences in temperature between treatments (Figure S1).
Pollen supplementation
To estimate pollen limitation in our experimental treatments, we hand-pollinated all flowers of HP females every other day with pollen from a pool of males from the same collection and kept in a greenhouse. For a given female and a given supplementation day, two pollen donors were randomly chosen in the pool and used to manually pollinate by brushing anthers of each donor on the stigmas (each male flower was used to pollinate on average five open female flowers).
Measuring floral traits
We measured a set of floral traits on all individuals in the experimental population, including traits that are presumably linked to pollinator attraction (corolla diameter, calyx height, flower number) and traits that are linked to individual fertility (gamete production per flower). Corolla diameter and calyx height were measured on two randomly chosen flowers per individual using a digital calliper precise to 0.01 mm. This was done twice, the first and last day of the experiment. Flower number was assessed on the same days. Measures for the 3 traits were averaged over the two dates prior to the analysis.
In addition, the number of gametes per flower was assessed for both sexes. For females, a total of 610 fruits (65% of the 933 fruits produced during the experiment) were dissected to estimate ovule production by imaging fruit content using a high resolution scanner (Epson Perfection V700 Photo,Seiko Epson, Suwa, Japan) and following the protocol described in Barbot et al. (2022).
For males, total pollen production per flower was estimated. To do so, one nearly opened flower bud per male was collected just before the experiment and dissected. Pollen quantity was assessed on two anthers using a particle counter (CASY® Model TT, Roche Innovatis AG, Bielefeld, Germany) and following the protocol described in Dufaÿ et al. (2008). Pollen quantity was then multiplied by five to get an estimate of pollen production per flower, since male Silene flowers have ten anthers.
Pollinator observations
Pollinator observations were conducted for each individual of the D group (i.e. diurnal pollination exposure treatment), using three 20 minutes sessions in the afternoon spread over the course of the experiment (for a total of pollination observations of 60 minutes per plant). Each pollinator visiting the experimental plot was visually identified (at the level of species, genus or family), and its sequence of flower visitation across plants was recorded. Two different variables were characterized per session: (i) the number of independent pollinators visiting a given plant and (ii) the total number of flowers visited per plant for all pollinators combined. Finally, we qualitatively characterized nocturnal pollinators diversity: observations were conducted for one hour the third day of the experiment, at 11pm, and involved (i) hanging a white sheet in the experimental garden and shining a mercury vapor lamp on it to attract, capture and identify nocturnal pollinators and (ii) direct observations on the plants using a flash-light.
Female reproductive success
During fructification, plants were surveyed twice a day in order to assess the prevalence of fruits predated by Hadena bicruris. Each time a caterpillar was recorded on a (primary) predated fruit, presumably shortly after emergence, we immediately removed it in order to avoid other (secondary) fruits predation events either on the same or a neighboring plant. In those primary predated fruits, all seeds are eaten (Jolivet & Bernasconi, 2006). We noted the number of (primary) predated fruits per female, which thus corresponded to the number of eggs laid by Hadena bicruris because females avoid already pollinated flowers for oviposition (Burkhardt et al., 2012; Castillo et al., 2013). It is noteworthy that this number underestimate predation effects on plant fitness, as caterpillars normally predate other fruits after emergence. Nonetheless, we employed this strategy to prevent the reproductive success of certain females from being compromised by their proximity to a predated plant in the greenhouse, as caterpillars can migrate between plants during the fruit ripening process.
Fruits were collected at maturity, typically three to five weeks after flower opening. Germination rates were estimated on a sub-sample of 60 seeds per female that were sown in petri dishes filled with 40mL of 10g/L agar in sterile water (photoperiod 14:10 and temperature 21-15°C). For each female, we estimated reproductive success as the number of viable seeds produced during the experiment, by multiplying mean seed number per fruit, total number of non-predated fruits and germination rate. The number of viable seeds was also computed using total fruit production instead of just non-predated fruits. Therefore, we have two estimators of female fitness, one that takes predation into account and one that does not. The number of dissected fruits per female to estimate seed and ovule number reflected its fruit production relatively to the overall fruit production at the population level (1 to 6 fruits dissected per female, mean = 4.68 ± 1,63 SD).
Male reproductive success
Male reproductive success was assessed by genotyping a subsample of seedlings (diurnal: 886 seedlings; nocturnal: 859 seedlings) and performing a paternity analysis. The number of genotyped seedlings per female reflected each mother’s contribution to the total seed pool, and seedlings were sampled randomly across the different fruits. Total genomic DNA from adults (120 females and 120 males) and seedlings was extracted and PCR assays were used to amplify five nuclear microsatellites following Barbot et al. (2022). Paternity analysis was then performed using a spatially explicit model derived from the mixed effect mating model (MEMM) developed by Oddou-Muratorio et al. (2018), as described in Barbot et al. (2022). For each male, this model computes Bayesian estimates of (i) mean pollen dispersal distance and (ii) reproductive success, defined as the sum, across all mothers, of the product of each putative mother reproductive success and the paternity share. Male reproductive success is thus an estimate of the number of seeds that a male sired across all seeds that were produced over the course of the experiment.
Mating success
To determine the number of reproductive partners for males and females, the same offspring were analysed using a likelihood-based paternity assignment approach, which allowed to identify the most likely father for each seedling and to reconstruct the father/mother/offspring trios (CERVUS v.3.0.7 software, Marshall et al., 1998; Kalinowski et al., 2007). For each plant, the mating success was the number of observed reproductive partners. These paternity analyses were conducted with an 80% confidence criterion and allowing for a 2% genotyping error.
Statistical analysis
Effect of the experimental treatments on the magnitude of pollen limitation and fruit predation
The occurrence of pollen limitation was tested on two components of female reproductive success by assessing differences between HP females and females from the four other groups (DC, DT, NC and NT) in (i) fruit-set (i.e. fruit number divided by the number of flowers produced during the experiment) and (ii) seed-set (i.e. seed number divided by ovule number, estimated on 1 to 6 fruits per female, see above). Regarding fruit-set, we conducted analyses using (i) primary fruit-set (including predated and non-predated fruits) and (ii) effective fruit-set (i.e. non-predated fruits only). The former index assesses whether there was enough pollen to maximize fruit initiation, while the latter index takes into account both benefit and potential cost of attracting pollinators. By definition, seed-set was estimated on non-predated fruits only. Differences among treatments in the two estimates of fruit-set were assessed using generalized linear models with a binomial error distribution, with a fixed effect of group (HP, DC, DT, NC and NT females), using the lme4 package (Bates et al., 2015). Differences in seed-set were assessed using a generalized linear mixed model with a binomial error distribution, with a fixed effect of group and a random effect of individual as several fruits were dissected per female. Post-hoc Tukey’s tests were performed to compare groups.
Finally, to assess the effect of treatments on predation, we conducted analyses on the proportion of the total number of fruits that had been predated over the course of the experiment, excluding HP females. To do so, we constructed a generalized linear model with exclusion and PAA treatment as explanatory variables with a binomial error distribution, and then proceeded to post-hoc Tukey’s tests to compare groups.
Diurnal pollinator activity
Generalized linear mixed models and type II ANOVA were used to investigate the effect of sex, PAA treatment and their interaction on both descriptors of pollinator visitation patterns (i.e. number of independent pollinators visiting a given plant and total number of visited flowers per plant) in plants exposed to the diurnal treatment (D). The four measured floral traits (i.e. flower number, corolla width, calyx height and gamete number per flower) were used as covariates, as well as their interaction with sex. Observation session and plant identity were fitted as random effects, as each individual was observed three times. The number of independent pollinators was analysed using a model with a Poisson error distribution while the total number of visited flowers per plant was analysed using a negative binomial distribution, to account for overdispersion and zero inflation. In the second model, the number of independent pollinators was added as covariate to assess, when one controls for the number of visiting pollinators, whether the treatment impacted the average number of flower visited per insect. These analyses were carried out both on the complete dataset and on a dataset comprising only plants visited at least once during the observation sessions. This last set of analyses was done in order to capture pollinator behavior (in terms of number of visited flowers) once they have arrived on a particular plant.
Effect of the experimental treatments on mating and reproductive success
As reproductive success was estimated using sex-specific estimation methods (i.e. seed counts for females and paternity analysis for males), the analyses presented below were performed independently on males and females. For males, we accounted for uncertainties in the estimation of male reproductive success by modeling prior weights of a posteriori distributions of the MEMM model (i.e., individuals with narrower confidence intervals in their estimation of reproductive success have less influence on the model).
Absolute reproductive success
To assess the effect of the treatments on individual fitness, we analyzed absolute reproductive success by using generalized linear models and type II ANOVA, with an effect of (i) exclusion treatment, (ii) PAA treatment and (iii) their interaction, as well as the four measured floral traits as covariates. We also compared variances in absolute reproductive success distribution between treatments using Levene’s tests. The same models were used to analyze variation in mating success (number of reproductive partners)
Selection gradients on floral traits
To compare the intensity of selection on floral traits among treatments, we performed analyses using relative reproductive success and standardized trait values, following standard recommendations (Lande & Arnold, 1983; De Lisle & Svensson, 2017). We computed relative reproductive success and standardized trait values per exclusion and PAA treatment, in order to compare selection gradients (i.e. the slopes of the regressions of reproductive success against phenotypic traits) between the four treatments. We first estimated selection gradients for each of the four PAA treatment x exclusion treatment combination, using multiple linear regression models with the four measured traits as independent variables as well as their interaction with the two treatments, by modifying contrasts and setting each treatment combination as base level. We then tested whether selection gradients differed between treatments by using type II ANCOVA.
Effect of the experimental treatments on pollen dispersal distance
We assessed whether mean pollen dispersal distances were affected by (i) the exclusion treatment, (ii) the PAA treatment and (iii) their interaction, using type II ANOVA. The four floral traits were also used as covariates in the model. Weights were included in the model to assess uncertainty of a posteriori individual mean pollen dispersal distances obtained with the Bayesian MEMM model.
Females and males with zero reproductive success (4 females and 8 males) were recorded as 0 for reproductive success analyses and as missing data for mating success analyses. DNA extractions having failed for seedlings from one maternal progeny, mating success is missing for an additional female. A few plants were excluded from the selection gradient analyses: one female plant due to a lost inflorescence, preventing flower production quantification, and five male plants due to missing pollen counts.
Results
No effect of treatments on pollen limitation but an increase in nocturnal fruit predation with PAA
The average number of fruits produced during the experiment was 11.22 (± SD 9.53), which represents about one-third of the usual annual fruit production for our collection of plants. Experimental treatments (HP DC, DT, NC and NT) did not affect neither seed set (\(\chi_{4,609}^{2} = 2.07,P = .73\)) nor primary fruit-set (\(\chi_{4,135}^{2} = 2.09,P = .72\)), suggesting pollen receipt did not limit female reproductive success in any of the experimental treatments (Table 1). Effective fruit-set (i.e. only non-predated fruits) significantly differed among experimental treatments (\(\chi_{4,135}^{2} = 19.23,P < .001\)). Tukey’s test revealed that it was significantly lower for NT females compared to DC females (\(P < .01\)) or DT females (\(P = .015\), Table 1), and lower also but not significantly so compared to NC females (\(P = .093\)) and HP females (\(P = .069\)). Overall, 5.1% of fruits produced during the experiment were predated, and this percentage increased to 10.6% for females only exposed to nocturnal pollinators, in particular in females with artificially increased PAA emission (5.94% in control females and 15.6% in females with increased PAA emission). When assessing the effect of treatment on the proportion of predated fruits, both exclusion treatment and PAA treatment had a significant effect on predation (\(\chi_{4,119}^{2} = 7.56,P < .01\) and \(\chi_{4,119}^{2} = 46.6,P < .001\) respectively). Tukey’s test revealed that proportion of predated fruits was significantly greater for NT females compared to DC females (\(P < .01\)), DT females (\(P < .01\)) and NC females (\(P = .034\), Table 1).
PAA did not significantly modify diurnal visitation patterns
In the experimental population, diurnal pollinators were mainly Bombus sp (B. terrestris, B. pascuorum and B. hortorum), as well as Syrphidae (Scaeva sp. and Sphaerophoria sp.). We also observed some occurrences of Halictidae (Lasioglossum sp.) and Sphingidae (Macroglossum stellatarum). Nocturnal pollinators included Noctuidae (Hadena bicruris and Autographa sp) and Pyralidae. Diurnal pollinators were more attracted by males than females, as indicated by the effect of sex on both the number of independent pollinators visiting a given plant (\(\chi_{1,345}^{2} = 36.6,P < .001\)) and the total number of flowers visited per insect (\(\chi_{1,345}^{2} = 11.02,P < .001\); Figure S2). PAA treatment did not significantly modify visitation patterns (independent visits: \(\chi_{1,345}^{2} = 0.86,P = .35\); total visits: \(\chi_{1,345}^{2} = 0.68,P = .41\)). Finally, the interaction term between PAA treatment and sex was not significant (independent visits: \(\chi_{1,345}^{2} = 0.053,P = .82\); total visits: \(\chi_{1,345}^{2} = 1.81,P = .18\)). Mean flower number and corolla width increased the number of independent pollinators attracted, while gamete number per flower decreased it (Table S1A). When analyzing the total number of visited flowers on individuals that were visited by at least one pollinator (i.e. by excluding the plants that were not visited during our sets of observation), we could also analyze in more details pollinators behavior once they arrived on a plant: we found that PAA addition increased the total number of visited flowers on the plant, but only in males (\(\chi_{1,82}^{2} = 5.22,P = .022\)). None of the traits showed a significant impact on the total number of visits at the plant level, whether non-visited plants were included or excluded (Table S1B).
Table 1 - Seed-set, primary fruit-set (including predated fruits) and effective fruit-set (only non-predated fruits) for each experimental female group. Significant or marginally significant differences in pairwise comparisons using Tukey’s test are indicated (a,b; \(P < .06\)).
Female group | Seed-set (± SE) | Primary fruit-set (± SE) | Effective fruit-set (± SE) | Proportion of predated fruits (± SE) |
HP (hand-pollinated) | 0.491 (± 0.024)a | 0.958 (± 0.017)a | 0.927 (± 0.021)ab | 0.031 (± 0.018)a |
DC (diurnal with control PAA) | 0.574 (± 0.017)a | 0.912 (± 0.027)a | 0.910 (± 0.027)a | 0.002 (± 0.002)a |
DT (diurnal with PAA addition) | 0.544 (± 0.019)a | 0.908 (± 0.029)a | 0.905 (± 0.038)a | 0.003 (± 0.003)a |
NC (nocturnal with control PAA) | 0.567 (± 0.018)a | 0.942 (± 0.020)a | 0.885 (± 0.061)ab | 0.067 (± 0.035)a |
NT (nocturnal with PAA addition) | 0.558 (± 0.019)a | 0.913 (± 0.028)a | 0.768 (± 0.021)b | 0.152 (± 0.006)b |
Table 2 - Absolute reproductive success (± SE) and mating success (± SE) per sex, per exclusion treatment (day D versus night N) and per PAA treatment (control C or manipulated T). Within sex significant differences in pairwise comparisons using Tukey’s test are indicated (a,b).
Reproductive success | Mating success | |||
Group | Female | Male | Female | Male |
DC (diurnal with control PAA) | 571.17 (± 72.33)a | 531.38 (± 83.82)a | 8.11 (± 0.76)a | 8.48 (± 0.65)a |
DT (diurnal with manipulated PAA) | 616.63 (± 81.19)a | 612.63 (± 82.72)a | 7.62 (± 0.75)a | 7.55 (± 0.68)ab |
NC (nocturnal with control PAA) | 561.76 (± 69.15)a | 561.94 (± 78.41)a | 6.86 (± 0.65)a | 6.81 (± 0.80)b |
NT (nocturnal with manipulated PAA) | 597.03 (± 114.25)a | 571.92 (± 68.53)a | 6.56 (± 0.59)a | 6.62 (± 0.65)ab |
No effect of exclusion and PAA treatments on reproductive success but an effect of exclusion treatment on mating success
Overall, PAA addition did not affect either female or male reproductive - RS - and mating success - MS - (females RS: \(F_{1,113} = 0.18,P = .68\); males RS: \(F_{1,114} = 0.20,P = .65\); females MS: \(F_{1,112} = 0.45,P = .50\); males MS: \(F_{1,106} = 1.68,P = .19\): ; Table 2). Similarly, the exclusion treatment did not affect reproductive success in either sex, meaning that seed production and siring success were comparable in plants exposed solely to diurnal pollinators versus nocturnal pollinators (females RS: \(F_{1,113} = 0.027,P = .87\); males RS: \(F_{1,114} = 0.039,P = .84\); Table 2). However, mating success was lower in both sexes for nocturnal pollination compared to diurnal pollination (females MS: \(F_{1,112} = 4.91,P = .027\); males MS: \(F_{1,106} = 9.98,P < .01\)). There was no significant effect of the interaction between PAA treatment and exclusion treatment (females RS: \(F_{1,113} = 0.0004,P = 0.98\); males RS: \(F_{1,114} = 0.39,P = 0.53\); females MS: \(F_{1,112} = 0.012,P = 0.91\); males MS: \(F_{1,106} = 1.4,P = 0.23\); Table 2). Finally, variances in reproductive and mating success were similar between groups as revealed by Levene’s tests (Table S2).
Table 3 - Results of the ANCOVAs analyzing female (left) and male (right) relative reproductive success in relation with the exclusion treatment, the PAA treatment and the four measured floral traits.
Variable | Females | Males | ||
F-value | P-value | F-value | P-value | |
Flower number | 31.54 | <.001 | 24.65 | <.001 |
Gamete number | 9.920 | .002 | 0.100 | .75 |
Corolla width | 0.008 | .93 | 1.470 | .30 |
Calyx height | 4.610 | .034 | 0.071 | .79 |
Exclusion | 0.046 | .83 | 0.046 | .83 |
Odour | 0.017 | .90 | 0.059 | .81 |
Flower number 🞨 Exclusion | 1.743 | .19 | 3.922 | .051 |
Gamete number 🞨 Exclusion | <0.001 | .99 | 0.183 | .67 |
Corolla width 🞨 Exclusion | 0.067 | .80 | 2.260 | .14 |
Calyx height 🞨 Exclusion | 1.428 | .24 | 0.940 | .34 |
Flower number 🞨 PAA | 0.057 | .81 | 2.252 | .14 |
Gamete number 🞨 PAA | 0.131 | .72 | 1.232 | .27 |
Corolla width 🞨 PAA | 1.564 | .21 | 0.002 | .96 |
Calyx height 🞨 PAA | 4.398 | .039 | 0.090 | .76 |
Exclusion 🞨 PAA | 0.080 | .78 | 0.228 | .63 |
Flower number 🞨 Exclusion 🞨 PAA | 0.297 | .59 | 4.857 | .03 |
Gamete number 🞨 Exclusion 🞨 PAA | 0.329 | .57 | 1.631 | .21 |
Corolla width 🞨 Exclusion 🞨 PAA | <0.001 | .99 | 3.419 | .068 |
Calyx height 🞨 Exclusion 🞨 PAA | 1.313 | .26 | 4.033 | .047 |
Notes: The table represents results from ANCOVAs for the main effect of floral traits and treatments, as well as two-ways and three-ways interactions between each variable. Statistics (F-values) and their associated P-values are indicated for each variable effect on female or male relative reproductive success.
Selection gradients differed between treatments
Males
Flower number was under positive selection in all groups of males except those exposed to diurnal pollinators and with no increased PAA emission, as shown by the significant three-way interaction term (Figure 1, Table 3 and Table S3). Pollen production per flower was not under selection in any of treatment combinations. We detected a significantly positive selection on corolla width only in males exposed to nocturnal pollinators and belonging to the control PAA group, along with a marginally significant effect of the three-way interaction term (Table 3). Finally, selection on calyx height was found to differ between groups (Table 3), although none of the selection gradients was significantly different from zero within each group (Figure 1 and Table S3).
Females
We found significantly positive selection on flower number in all treatment combinations (Figure 1). Gamete number was under statistically significant positive selection only in PAA females exposed during the day, although the three way interaction was not significant (Table 3). Corolla width was not associated with increased female reproductive success in any of the four groups. Concerning calyx height, we found significant selection on this trait in females pollinated during the night and with increased PAA emission, but not in the other groups. The impact of treatments on selection on this trait was slightly different when looking at interactions: three way interaction was not significant, whereas significant calyx height x PAA interaction suggests a stronger selection on the trait for all female with increased PAA emission (Table 3). All these results were identical using total fruit production instead of non-predated fruits in the estimation of female reproductive success (Figure S4, Table S4).
Nocturnal pollinators dispersed pollen further
Mean pollen dispersal distance was higher during the night (diurnal males: 1.62m ± 0.032, nocturnal males: 1.98m ± 0.074; \(F_{1,115} = 4.62,P < .001\); Figure S3) but was not affected by PAA treatment (\(F_{1,115} = 0.42,P = .16\)). There was no effect of the interaction between exclusion and PAA treatments on pollination distances (\(F_{1,115} = 0.0054,P = .87\)).
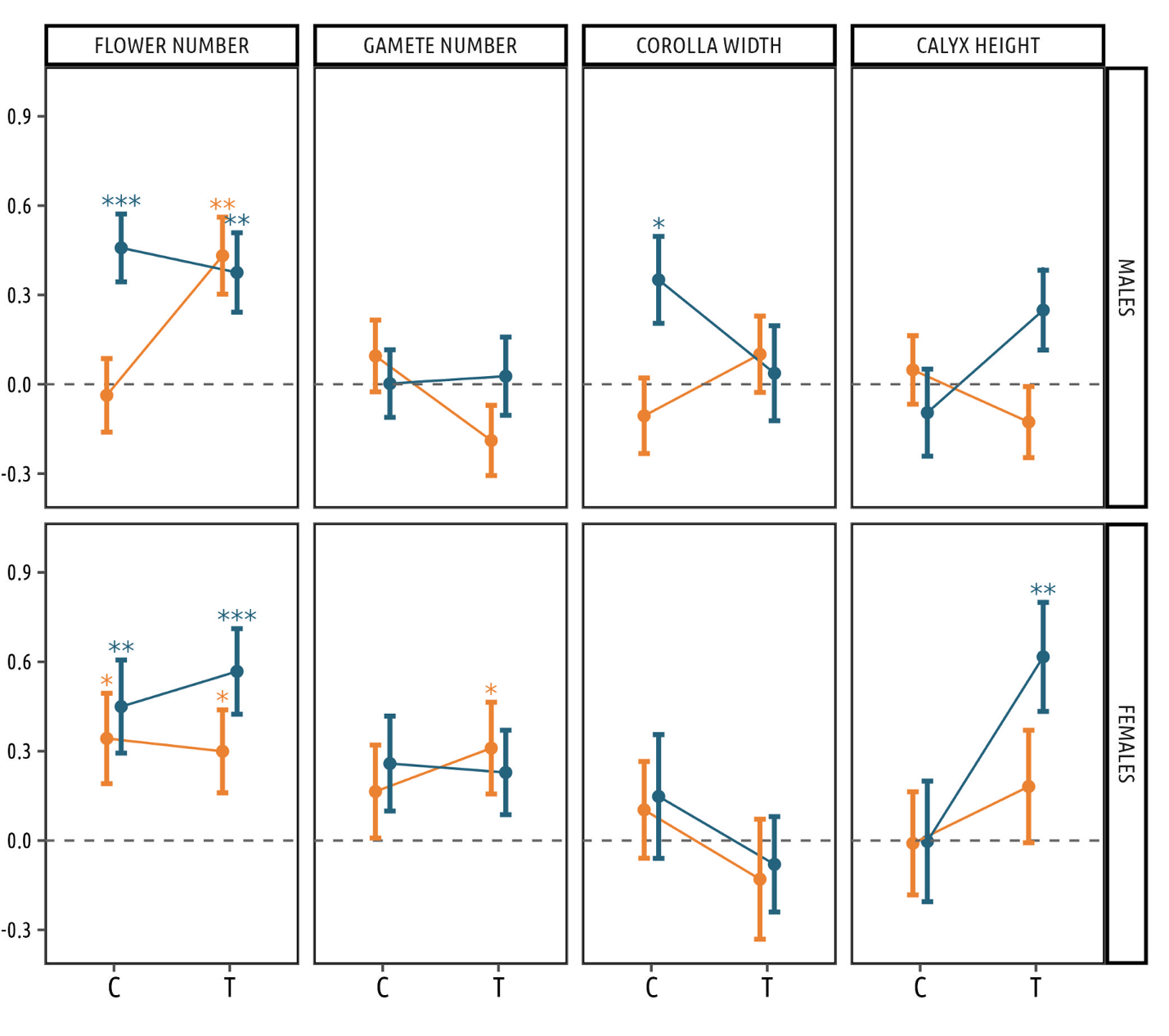
Figure 1 - Estimates of selection gradients (± SE) on floral traits in each sex, and for the four treatment combinations. Yellow dots and error bars represent selection gradient estimates for the Diurnal pollination group (D), whereas blue dots and error bars represent selection gradient estimates for the Nocturnal pollination group (N). Within each panel, selection gradient estimates for the PAA control group (C) are represented on the left, whereas selection gradient estimates for the PAA manipulated group (T) are represented on the right. P-values are represented with * : P < 0.05, ** : P < 0.01 and *** : P < 0.001. Asterisks on the top of error bars refer to selection gradients that are significantly different from zero in that particular group (i.e. PAA treatment x exclusion treatment combination).
Discussion
A true mixed pollination system: an opportunity to study pollinator-mediated selection
In this study, by comparing groups of plants pollinated during day versus night, we investigated how pollinator groups shape pollen dispersal distances, access to sexual partners and reproductive success in Silene dioica. This species is generally considered to have a generalist diurnal pollination system, although it is also visited by nocturnal pollinators (Jürgens et al., 1996; Kephart et al., 2006). Here, we not only observed pollination in both groups of plants exposed to diurnal and nocturnal insects but also uncovered a genuine mixed pollination system. Indeed, we found no evidence of pollen limitation during either day or night, and there were no discernible differences in male or female reproductive success between exposure treatments. These findings underscore once again the unreliability of pollination syndromes as predictors of pollination regimes in Silene species (Prieto-Benítez et al., 2015).
In a species with a true mixed pollination system, diurnal versus nocturnal pollinators exclusion experiments offer the opportunity to dissect the components of overall selection and to compare groups of pollinators. Previous studies on Silene dioica have found that selective pressures acting on floral traits in females, such as flower and ovule number, were not mediated by pollinators, but were rather linked to fecundity selection (Barbot et al., 2022, 2023). While investigating the nature of selection (pollinator-mediated versus fecundity selection) is possible in female plants by comparing selection gradients between open- and hand-pollinated plants (Caruso et al., 2019), this cannot be done in male plants. Males of Silene dioica are known to be under positive selection for flowering duration and flower size (Barbot et al., 2023), but direct evidence for the role of pollinators in these selective pressures is currently lacking. Our manipulative approach forced us to reduce our study to a relatively short period of time compared to the length of flowering season in S. dioica. Evidently, we could not, with this study, confirm the positive selection acting on flowering duration in males. However, the fact that a large part of selection gradients documented here were consistent with those estimated throughout the whole flowering season in the same species (Barbot et al., 2023) suggests that our measurements are likely to be representative of what happens over a complete reproductive episode. As discussed below, females were found to be selected to produce more flowers and more ovules, whereas selection toward larger flowers was found in males. The confrontation of these gradients during day versus night now allows us to better understand the exact mechanisms involved. Here, we compared diurnal versus nocturnal components of selection in males, and any difference detected in selection gradients can be attributed to differences in pollinators behavior and/or abundances between treatments, thus indirectly supporting the notion of pollinator-mediated selection in males.
Nocturnal pollinators disperse pollen further, but reach less sexual partners
One notable distinction observed between plants exposed during the day versus night pertained to pollen dispersal. Our data supports extremely limited pollen dispersal distances, a characteristic often observed among herbaceous plants (De Cauwer et al., 2012; Tonnabel et al., 2019), particularly in insect-pollinated species (Hardy et al., 2004; Llaurens et al., 2008; Van Rossum et al., 2011). In our study, nocturnal pollinators dispersed pollen on average 18.5% further than diurnal ones. This effect has also been observed in Silene latifolia either by using fluorescent powders to assess pollen dispersal distances (Shykoff & Bucheli, 1995; Young, 2002), or by genotyping seedlings as done our study and in Barthelmess et al. (2006). Other studies on Oenothera harringtonii demonstrated that hawkmoths, the primary pollinators in this system, facilitate long-distance pollen dispersal thereby reducing genetic isolation through gene flow (Rhodes et al., 2017; Skogen et al., 2019). In parallel, increased pollen dispersal distances during the night led to decreased mating success in both sexes. This outcome may be attributed to the generally higher abundance of diurnal pollinators compared to nocturnal pollinators (Knop et al., 2018; Zoller et al., 2020; Anderson et al., 2023), a trend that has also been observed in the sister species S. latifolia (Altizer et al., 1998). Differences in pollinator foraging behavior and/or in carry-over could also explain why nocturnal pollination leads to lower mating success despite increased pollen dispersal distances (Miyake & Yahara, 1998; Castellanos et al., 2003). In any case, disparities in both pollen dispersal distance and mating success between nocturnal and diurnal pollinators could strongly impact spatial genetic structure in natural populations as well as the exact scale on which selection processes on floral traits operate (Brunet et al., 2012; Gamba & Muchhala, 2020).
Artificially increasing PAA emission results in higher fruit predation by nocturnal pollinators
One aim of this study was to assess whether variation in one type of signal (volatile compound) could influence patterns of selection on other floral characteristics (visual signals). To test this, we exaggerated PAA emission in half of the plants, a compound known to be abundant in the floral bouquet of S. dioica and important for pollinator attraction in many systems, with either diurnal or nocturnal pollination (Cantelo & Jacobson, 1979; Heath et al., 1992; Tóth et al., 2010). Clearly, this is only a first step in understanding the interplay between scent emission and selection on other attractive traits, as the caricatural variation in PAA used here does not reflect the quantitative variation that is likely to occur in natural populations. A more detailed understanding of selection on VOC emissions — including PAA — and the interaction between selection on scent signals and visual signals will require characterizing the floral bouquet and quantifying the strength and direction of selection acting on these traits. While we found no increase in female reproductive success, we observed a rise in fruit predation rate associated with PAA addition. This phenomenon may be attributed to a positive effect of PAA emission on Hadena's visitation without any benefit in seed production, because high pollinator abundances already saturated female reproductive success. Another non-exclusive explanation would be that PAA triggered oviposition behaviour through chemotactile receptors of the ovipositor female moths (Dötterl et al., 2009). This impact of the PAA treatment on predation intensity appears to influence selective pressures on certain floral traits, a topic that will be further discussed below.
Floral traits are mostly under fertility selection in females
The absence of pollen limitation in all experimental groups implies a lack of pollinator-mediated selection in females under our experimental conditions. Accordingly, most selection gradients were found to be similar across pollination treatments. The positive selection on flower number in all treatments suggests that the pollinator group (diurnal versus nocturnal) does not significantly influence this pattern. In other terms, selection on flower number could only be ascribed to fertility selection: females that produce many flowers have a better fitness because they produce more gametes, and not because they attract more pollinators. These results align with findings from previous studies on the same species (Barbot et al., 2022, 2023), and further underscore that the pollinator group (night versus day) generally does not exert a significant influence on the patterns of selection on floral traits in females.
A notable exception was observed regarding calyx height, which was found to be under positive selection but only in plants exposed during the night and with an artificial increase of PAA. This result could be associated with the observed rise in fruit predation within this particular group of plants. Nonetheless, calyx height has been reported to be positively associated with likelihood of oviposition by Hadena sp in other Caryophyllaceae species (Kula et al., 2013; Miyake et al., 2018), so the proximal cause of this pattern remains undetermined.
Pollinator-mediated selection on floral traits in males
In males, we observed differences in selection patterns among pollination treatments, implying that pollinator behavior and visitation patterns play a stronger role on fitness compared to females. These results align with theory, as males should be more limited in their reproductive success by access to sexual partners and/or gametes through pollinator attraction than females (Arnold, 1994; Delph & Ashman, 2006; Moore & Pannell, 2011). This observation also implies that nocturnal and diurnal pollinators mediate contrasting selective pressures on male S. dioica, consistent with findings in other systems (Young, 2002; Kulbaba & Worley, 2012; Scopece et al., 2018). When focusing on control plants, we indeed found that only males exposed to nocturnal insects experienced positive selection on flower number and corolla width. One logical explanation, although not directly observed in the current study, would be that nocturnal insects prefer to visit and/or spend more time on plants with large flower numbers and large corollas. This is consistent with a previous study conducted on two Silene species, S. latifolia and S. diclinis, which showed that large flowers were more likely to be predated by Hadena bicruris (Brothers & Atwell, 2014). Because Hadena bicruris does not discriminate between sexes when visiting the sister species Silene latifolia (Labouche & Bernasconi, 2009), this could result in males with larger floral display being more frequently visited by nocturnal pollinators.
A previous study conducted on Silene dioica found positive selection on corolla width in males exposed to both pollinator categories, without exclusion experiments (Barbot et al., 2023). While this was interpreted as the consequence of the observed preference of Bombus terrestris for large flowers (Moquet et al., 2022) it now appears that such selection patterns may be mainly driven by floral traits preferences of nocturnal pollinators. Regarding selection on flower number, our current results contrast with selection patterns detected in previous studies, which documented no selection on flower number in males when plants are visited by mix of diurnal and nocturnal pollinators (Barbot et al., 2023). Studies of Bombus terrestris behavior showed that flower number in Silene dioica is a strong signal increasing the number of pollinator visits (Moquet et al., 2022). Unexpectedly, when focusing on selection patterns during the day, we found that the attractive function of flower number did not translate into positive selection on this trait. This probably decreases the overall benefit of producing high numbers of flowers even though nocturnal pollinators mediate some positive selection on this trait.
Interestingly, these differences in selective patterns between plants visited by nocturnal versus diurnal insects appear to interact with the experimental increase in PAA emission. In the group of males exposed to nocturnal pollinators, the PAA treatment modified the observed selective pressures, by removing selection on corolla width and leading to a significantly stronger selection in favor of longer calyxes. If, as in the sister species Silene latifolia (Dötterl et al., 2005), the majority of benzenoids, including PAA, are released by the petals in S. dioica, then variation in corolla diameter among individuals may result in differences in natural scent emission levels. The selection pressure exerted by nocturnal pollinators on corolla width could thus diminish with the exacerbation of PAA emission, as pollinators may fail to detect olfactory differences in corolla size among plants. Moreover, in males exposed during the day, the treatment led to a positive selection on flower number. Since we observed that insects, once arrived on the males with increased PAA emission, stayed longer (i.e. visited more flowers), the PAA may have lead insects to more efficiently remove (and export) pollen grains, for a given value of flower number, which may in turn translate in a positive selective pressure acting on this trait. It thus appears that the artificial increase in PAA emission interacts with other traits involved in pollinator attraction, thereby modifying the identity of plant attractive signals for pollinators, or the sensibility of the latter to these attractive traits (Fenster et al., 2015). Nonetheless, given that the artificial increase of PAA may not accurately reflect natural conditions, future studies should focus on measuring selection gradients for individual compounds involved in scent emission. This would allow for a more accurate assessment of the selection gradient on PAA emission and its interaction with the selection of other floral traits.
Conclusion
In conclusion, this study revealed more pronounced responses of selection gradients on floral traits to changes in groups of pollinators and in the emission level of a volatile organic compound in males compared to females. This global pattern could be interpreted as indirect evidence that males are more dependent of pollinator attraction than females in their reproductive success. Although theory indeed predicts that male function should depend more on pollinators compared to females, such prediction is usually difficult to verify empirically. Experimental manipulation of pollinator identity combined with the measurement of selection gradients thereby offers a promising approach for studying patterns of pollinator-mediated selection.
Acknowledgements
We would like particularly to thank E. Klein for letting us access the MEMM model, and J. Tonnabel and M. Dubart for their advices about MEMM model implementation. We thank S. P. De Lisle for precious advices about reproductive and mating success relativization and traits standardization in selection gradient analysis. For laboratory assistance, we thank L. Debacker, S. Flourez and N. Le Roux. This work has been performed using infrastructure and technical support of E. Schmitt and N. Faure from the Plateforme Serre, cultures et terrains expérimentaux – Université de Lille for the greenhouse/field facilities. Preprint version 3 of this article has been peer-reviewed and recommended by Peer Community In Evolutionary Biology (https://doi.org/10.24072/pci.evolbiol.100798; Arroyo & Simón-Porcar, 2024).
Funding
The authors declare that they have received no specific funding for this study.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data, scripts, code, and supplementary information availability
Data are available online: https://doi.org/10.5281/zenodo.11487468 (Barbot 2024a). Scripts and code are available online: https://doi.org/10.5281/zenodo.11487700 (Barbot 2024b). Supplementary information is available online: https://doi.org/10.5281/zenodo.11487621 (Barbot 2024c).