
 CC-BY 4.0
CC-BY 4.0
Introduction
Since its registration as a broad-spectrum contact herbicide in 1974, the use of glyphosate (GLY) has continuously increased, making GLY-based pesticides the most widely applied herbicides worldwide (Richmond, 2018; Gandhi et al., 2021). Maggi et al. (2020) reported that GLY and its main degradation product aminomethylphosphonic acid (AMPA) may be persistent contaminants in approximately 30% and 93% of global cropland soils, respectively. On the basis of persistence, accumulation and toxicity criteria, they computed that 1% of croplands worldwide (i.e. approximately 385,000 km2) experience mid-high to high hazard conditions. Outside these hotspots, contamination is expected to be low but globally pervasive (Maggi et al., 2020).
Initially, GLY was marketed as an “environmentally friendly” herbicide owing to its low expected environmental persistence (easily biodegraded), low predicted toxicity to wildlife and humans (absence of the shikimate pathway in animals, which is found only in plants and microorganisms and is the target pathway of GLY in plants), and lack of potential for bioaccumulation (hydrophilic, low log octanol–water partition coefficient Kow = − 3.12), poorly metabolized and rapidly eliminated in mammals) (EFSA, 2015a; Kissane & Shephard, 2017; Richmond, 2018; Gandhi et al., 2021). The log Kow is a measure to assess whether a substance is more or less hydrophobic (and/or lipophilic, fat solubility) or hydrophilic (and/or lipophobic, water solubility). A high bioaccumulation potential may be expected for log Kow > 3, and Kow is used as one of the regulatory criteria to classify a compound as potentially bioaccumulative if log Kow > 4.5. However, concerns about the environmental safety and health risks associated with GLY have increased within recent decades, especially in the context of its continuously increasing use (Vandenberg et al., 2017; Székács & Darvas, 2018; Van Bruggen et al., 2018; Gandhi et al., 2021). GLY and AMPA are more persistent in the environment than initially assessed, as shown by the ubiquitous occurrence of residues in soil, water, and air (Bai & Ogbourne, 2016; Richmond, 2018; Van Bruggen et al., 2018; Xu et al., 2019; Gandhi et al., 2021). They can accumulate in environmental compartments, crops, and the human food chain (Bai & Ogbourne, 2016; Richmond, 2018). They are associated with chronic low-dose exposure risks in animals and humans, and can be more harmful to ecosystems than expected (Kissane & Shephard, 2017; Richmond, 2018; Peillex & Pelletier, 2020; Gandhi et al., 2021; Ojelade et al., 2022). Furthermore, GLY and AMPA may be more bioaccumulative substances than predicted from their physico-chemical properties (Richmond, 2018; Ruuskanen et al., 2020; Serra et al., 2021; Pelosi et al., 2022). However, the controversial issues regarding GLY-based herbicides have focused mostly on human health (Vandenberg et al., 2017; Székács & Darvas, 2018), whereas GLY and AMPA exposure and their impacts on wildlife have been overlooked (Kissane & Shephard, 2017).
In the USA and EU, registration reviews concerning GLY use as a plant protection product (PPP) identified potential risks to terrestrial and aquatic plants and nontarget terrestrial arthropods, and risks to mammals and birds (EFSA, 2015a; 2015b; 2015c; US EPA, 2020). In the EU assessment for vertebrates, the taxa identified as the most at risk were mammals, particularly herbivorous mammals (EFSA, 2015a; 2015b). Glufosinate (GLUF), like GLY, is an organophosphorus compound used as a broad-spectrum nonselective contact herbicide with a systemic action and was considered “one of the very few alternatives to glyphosate” (European Commission, 2017). However, risks for herbivorous and insectivorous mammals associated with the herbicidal use of GLUF have been reported in environmental risk assessment (ERA) processes (EFSA, 2005, 2012). GLUF was classified as a candidate for substitution in the EU according to Commission Regulation EC No 1107/2009 (PPP Regulation) in 2015, and withdrawal was scheduled at its end of approval in the EU in 2018 (Robin and Marchand, 2023). However, GLUF is still used in many countries worldwide.
The singular physicochemical properties of GLY (a particularly hydrophilic polar compound, existing in different ionic states depending on pH, making it difficult to extract) and closely related compounds such as AMPA or GLUF make measurement of their residues challenging and prevent these compounds from being included in multiresidue analyses (Delhomme et al., 2021). Such technical limitations in analytical chemistry may have hampered the development of studies about GLY, AMPA and GLUF exposure and their effects in wildlife and, more generally, knowledge building about the terrestrial ecotoxicology of these compounds. Ultimately, this lack of knowledge could be an impediment for regulatory decisions with respect to the objectives of reducing the risks and impacts of pesticide use on human health and the environment (e.g. Directive 2009/128/EC). Assessing and characterizing the exposure of wildlife to GLY, AMPA and GLUF is therefore a critical issue for environmental protection and biodiversity conservation.
The main aim of this study was to investigate the exposure of wildlife in an arable landscape to GLY, AMPA and GLUF and evaluate the risk for toxicological impairment, with a focus on small mammals. This was achieved via a minimally invasive sampling method based on residue analyses in hair and on reverse dosimetry approaches. The compound residues can be detected and quantified in hair, owing to the effectiveness of this matrix for assessing exposure to multiple pesticides in wildlife (Krief et al., 2017; Fritsch et al., 2022), as shown for GLY in bats (Hooper et al., 2022) and both GLY and AMPA in humans (Alvarez et al., 2022). Moreover, GLY concentrations were relative to the intensity of exposure in human samples (i.e. GLY was quantified at lower concentrations in hair samples from nonoccupationally exposed subjects than in hair samples from farmers, who often used GLY) (Alvarez et al., 2022).
Differences in exposure between the three compounds were investigated, and the influences of farming practices and herbicide use intensity were studied. We expect exposure to GLY to be greater than that to the other compounds. Mammals can be exposed to herbicides through diet and overspray (European Commission, 2015; Martinez-Haro et al., 2022; Mayer et al., 2020; Newton et al., 1984). GLY is more commonly used than GLUF, and GLUF use is not longer authorized in France (BNV-D Traçabilité). AMPA is not a pesticide active substance itself but a metabolite of GLY. Therefore, while small mammals may be exposed to the three compounds through trophic exposure, they could be additionnaly exposed through overspray only in the case of GLY. Higher concentrations in hair were expected in treated areas (i.e. conventional crops) than in nontargeted habitats (i.e. organic farming (OF) fields and hedgerows). Since small mammals are mobile species, the influence of herbicide use intensity was also considered at the landscape scale (i.e. using the proportion of OF fields within the landscape as a proxy) and at the township level (i.e. using sales of active substances as a proxy of usage intensity). To further document the factors that may shape exposure to GLY, AMPA and GLUF, differences between small mammal species (rodents and shrews) with various ecological traits were investigated. Herbivorous voles are expected to exhibit greater exposure than are granivorous, omnivorous or insectivorous species according to the higher daily dietary doses estimated during ERA processes (EFSA, 2005, 2012; European Commission, 2015). Moreover, herbivorous voles are less mobile than omnivorous mice and voles or shrews are (van den Brink et al., 2011; Wijnhoven et al., 2006) and may not avoid treated areas. We expect species that feed on animal matter (i.e. omnivores, carnivores, insectivores or vermivores) to also be exposed. Indeed, GLY residues have been found in small carnivorous and omnivorous mammals in a forest ecosystem (Newton et al., 1984), and residues of GLY, AMPA and GLUF have been found in earthworms collected from the study area, a taxa that can be prey of rodents and shrews (Pelosi et al., 2022).
Material and methods
Study site and design
The study site was the Long-Term Socio-Ecological Research (LTSER) “Zone Atelier Plaine & Val de Sèvre” (ZAPVS, https://za-plaineetvaldesevre.com/) in mid-western France (see Bretagnolle et al., (2018) for a general site description). The study was conducted within the framework of a larger project which acronym was “PING”, for which the sampling design was based on about 40 sampling squares of 1 km2, selected on the basis of landscape features (gradient of seminatural habitats such as hedgerows and a gradient of organic farming surfaces), in which sampling was conducted in cropped and semi-natural habitats (cf. Pelosi et al., 2022). The trapping of small mammals was conducted in the 1 km2 sampling squares in both an arable cereal field (winter cereals) and a woody patch (hedgerow or woodlot edge) as close as possible to the sampled arable field (Fig. 1). Cropped cereals and hedgerows are referred to as “habitats” hereafter. Cropped fields were cultivated either under conventional farming (CF) or organic farming (OF). The farming practices in the organic fields followed the rules of the “AB (Agriculture Biologique) France label” and were under OF for at least three years at the time of sampling. A few fields were in transition from CF to OF and were referred to as “OF/CF”.
Sampling process and sample size
Sampling was conducted in spring 2018 (May-June). Small mammals were captured using traplines of nonlethal traps (25 traps, 2 m apart) with dormitory (Le Quilliec & Croci, 2006) filled with hay and food (i.e. pieces of carrots and apples, sunflower seeds, cat food and peanut butter). The hay, vegetables, fruits and seeds originated from organic agriculture. The traps were checked every day, rebaited and refilled or replaced as necessary. The traplines were set during one to four nights depending on logistics and capture success (i.e. longer duration, if possible, in case of an absence of capture during the first night(s)). Species were determined during the handling of individuals on the basis of morphometrics and confirmed by molecular analyses in case of doubt. Microtus arvalis and Microtus agrestis being morphologically similar species, the identification of Microtus voles was confirmed by molecular analyses (Sup Info Annex 1. Text A1). The suspected Mus spp individuals were also confirmed genetically. For that, a tiny piece of ear was sampled (using medical clean and disinfected scissors, followed by disinfection of the ear) and stored by freezing (-20 °C) until DNA extraction. A sample of 50-100 mg of hair was taken by shaving, wrapped in aluminium foil and stored in plastic zip-lock bags at -20 °C until analysis. Individuals were released after handling. The survival rate to capture and handling was 91.6%. In case of recapture, the individuals were immediately released without any measurement or sampling (recapture rate around 3%).
The experiments were performed under the authorization of the French National Ethical Committee (Project APAFIS N°5340) by skilled and experienced investigators from the Chrono-environnement research department (EU0592), following directive 2010/63/EU on the protection of animals used for scientific purposes. All precautions to limit stress and deleterious effects on the animals as much as possible were taken. The sampling and handling of small mammals were conducted under supervision and with the participation of people authorized for animal experimentation and experienced in small mammal capture and handling and using appropriate and authorized (EU0592) facilities (a vehicle equipped with a mobile anaesthesia unit and all of the materials and equipment required for animal welfare).
The sample size resulted in hair samples obtained from 61 individuals captured in 29 traplines distributed over 25 squares of 1 km2 (Fig. 1, Table 1; further details about the sample sizes are provided in Sup Info Annex 1. Table A1). Individual samples of 18 wood mice (Apodemus sylvaticus), 14 greater white-toothed shrews (Crocidura russula), 16 common voles (Microtus arvalis), 11 bank voles (Myodes glareolus), and two house mice (Mus musculus) were analysed. A total of 41 samples were collected from individuals captured in plots from CF farming category, 10 from OF/CF category, and 10 from the OF category. A total of 39 samples were collected from hedgerows or woodlots, and 22 were collected from cereal fields. Forty-one samples came from habitats classified as non-targeted by PPP treatments and 20 from plots classified as targeted by PPP treatments.
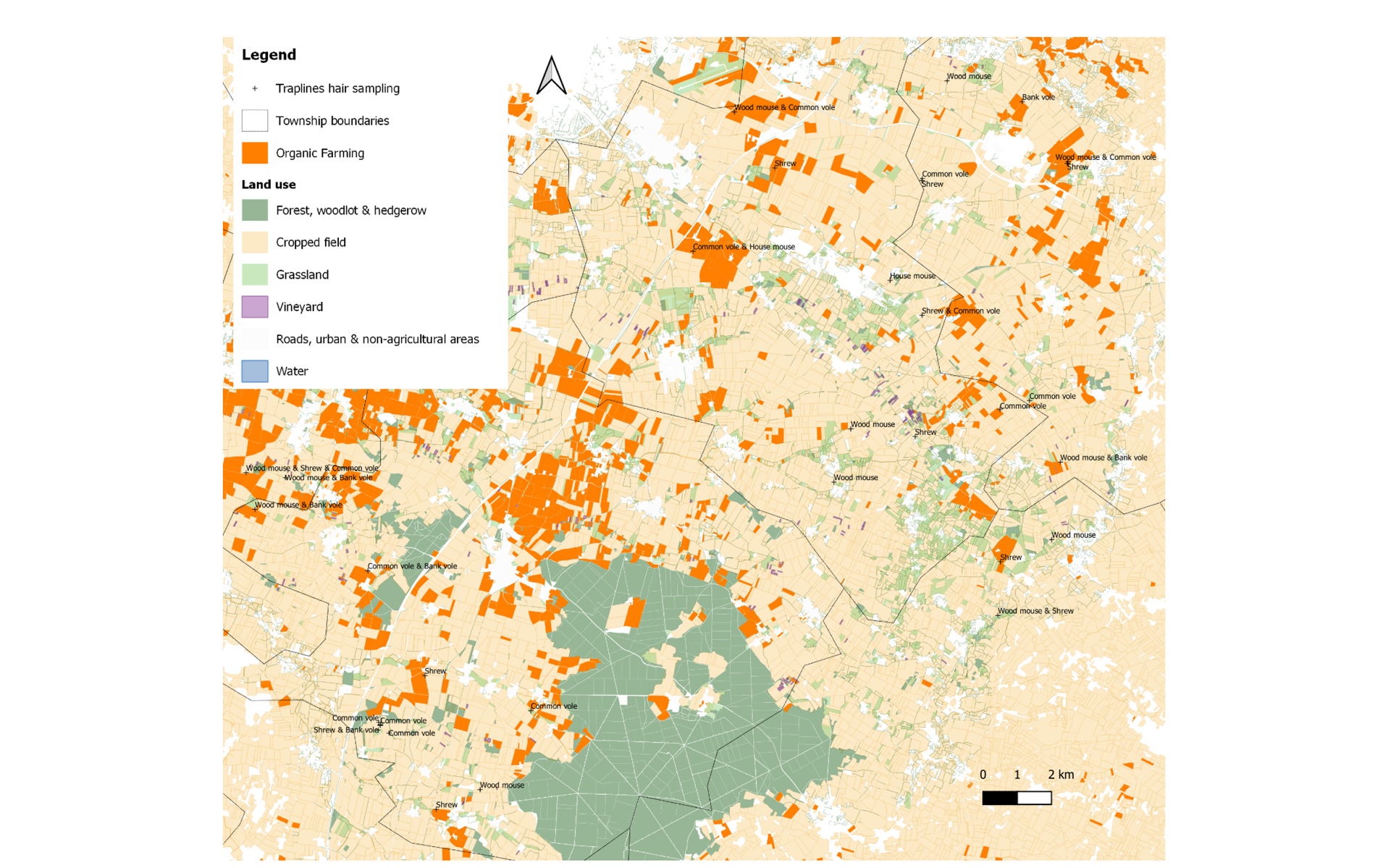
Figure 1 - Map of the traplines where hair samples were collected across the study area.
Table 1 - Sample size and frequency of detection and concentrations of glyphosate, AMPA and glufosinate in the hair of small mammals. Descriptive statistics on concentrations were computed when the given compound was detected, i.e. nondetects (samples whose concentrations were below the lowest detected value (Minimum)) were excluded. Home range size were obtained from Chambers et al. (2000) and Wijnhoven et al. (2006). The bank vole is often considered a granivorous species but recent studies have shown omnivory. Abbreviations: OF=Organic farming, CF=Conventional farming, PPP=plant protection product, NT=Not targeted by treatments, T=Targeted by treatments, LOQ= Limit of quantification.
All species | Wood mouse Apodemus sylvaticus | Greater white-toothed shrew Crocidura russula | Common vole Microtus arvalis | House mouse Mus musculus | Bank vole Myodes glareolus | |
|---|---|---|---|---|---|---|
Diet | Omnivore | Insectivore | Herbivore | Omnivore | Omnivore | |
Mean home-range size (m2) | 230–12200 | 50–200 | 161–1350 | 2710 | 260–4100 | |
NUMBER OF INDIVIDUALS | ||||||
Habitat | ||||||
Cereal | 22 | 9 | 2 | 9 | 2 | |
Hedgerow or woodlot | 39 | 9 | 12 | 7 | 11 | |
Farming | ||||||
CF | 41 | 12 | 9 | 9 | 1 | 10 |
OF/CF | 10 | 4 | 3 | 2 | 1 | |
OF | 10 | 2 | 2 | 5 | 1 | |
Targeted or not by PPP treatments | ||||||
« NT » | 41 | 9 | 12 | 8 | 1 | 11 |
« T » | 20 | 9 | 2 | 8 | 1 | |
Landscape OF proportion | ||||||
0-50% | 40 | 11 | 9 | 8 | 1 | 11 |
50-100% | 21 | 7 | 5 | 8 | 1 | |
Sales GLY (kg) | ||||||
< 7000 | 25 | 6 | 4 | 11 | 2 | 2 |
> 7000 | 36 | 12 | 10 | 5 | 9 | |
Sales GLUF (kg) | ||||||
0 | 44 | 15 | 11 | 7 | 11 | |
0.75 | 17 | 3 | 3 | 9 | 2 | |
Total | 61 | 18 | 14 | 16 | 2 | 11 |
GLYPHOSATE | ||||||
Detections | ||||||
Number of detection | 39 | 6 | 7 | 15 | 1 | 10 |
Frequency (%) | 64 | 33 | 50 | 94 | 50 | 91 |
Concentrations (pg/mg) | ||||||
LOQ | 5 | |||||
Minimum | 0.018 | 0.600 | 0.600 | 0.018 | 1.43 | |
Q1 | 1.040 | 0.795 | 0.600 | 2.23 | 2.60 | |
Median | 2.65 | 1.77 | 0.800 | 4.42 | 3.79 | 3.65 |
Q3 | 5.77 | 4.83 | 0.950 | 8.13 | 5.63 | |
Maximum | 522 | 5.92 | 1.22 | 522 | 7.74 | |
AMPA | ||||||
Detections | ||||||
Number of detection | 31 | 4 | 2 | 14 | 1 | 10 |
Frequency (%) | 51 | 22 | 14 | 88 | 50 | 91 |
Concentrations (pg/mg) | ||||||
LOQ | 1 | |||||
Minimum | 0.240 | 0.400 | 0.400 | 0.240 | 0.515 | |
Q1 | 1.025 | 0.519 | 0.617 | 1.08 | 1.19 | |
Median | 1.39 | 1.29 | 0.833 | 2.51 | 1.37 | 1.61 |
Q3 | 6.45 | 2.03 | 1.05 | 7.65 | 6.51 | |
Maximum | 33.6 | 2.07 | 1.27 | 19.4 | 33.6 | |
GLUFOSINATE | ||||||
Detections | ||||||
Number of detection | 27 | 2 | 1 | 13 | 1 | 10 |
Frequency (%) | 44 | 11 | 7 | 81 | 50 | 91 |
Concentrations (pg/mg) | ||||||
LOQ | 1 | |||||
Minimum | 1.16 | 2.04 | 1.29 | 1.16 | ||
Q1 | 2.01 | 7.88 | 2.39 | 1.59 | ||
Median | 3.51 | 13.8 | 1.48 | 4.05 | 9.75 | 3.01 |
Q3 | 7.25 | 19.7 | 5.80 | 4.87 | ||
Maximum | 25.5 | 25.5 | 21.8 | 12.6 |
Proxies related to the intensity of treatments at various spatial scales
First, to investigate the role of agricultural practices in shaping the exposure of small mammals, habitats were classified as cultivated under OF or CF according to the type of agricultural organic or conventional practices in the cultivated fields. To classify hedgerows or woodlots with respect to farming practices, the adjacent plots were considered, and each woody patch was classified as “OF” if adjacent fields were under OF and none of the adjacent fields was under CF, “OF/CF” if at least one of the adjacent fields was under CF, or “CF” when all adjacent fields were under CF.
For the proxies of treatment intensity, at the plot scale (arable field or woody patches where trapping was realized), a general proxy of potential for pesticide treatment was computed. The fields cultivated under OF and hedgerows or woodlots were considered not intentionally targeted by treatments (NT), whereas the arable fields cultivated under CF and transitional fields cultivated from CF to OF were considered possibly targeted (T).
A second proxy to characterize the intensity of potential treatments was computed at the landscape scale. The locations and boundaries of cultivated plots in 2018 were obtained from the official French “Registre Parcellaire Graphique” (https://www.data.gouv.fr/fr/datasets/parcelles-en-agriculture-biologique-ab-declarees-a-la-pac). The software Quantum GIS (QGIS 3.16) was used to calculate the proportion of OF plots in the landscape, which was computed in each sampling square of 1 km2 as the proportion of the total surface occupied by OF arable fields. The final proxy resulted in a two-level factor, with a category “OF dominant” with proportions varying from 50 to 100% ([49-79], 6 sampling squares), in which treatment intensity at the landscape scale was expected to be lower since arable fields not targeted by pesticide treatments prevailed within the landscape mosaic. The second category, “CF dominant”, gathered squares with landscape OF proportions ranging from 0 to 50% ([0-49[, 19 sampling squares).
Finally, to consider the intensity of potential treatments over the range of several farmlands, a proxy at the township scale was computed. The sales of active substance (a.s., data provided in kg) were obtained from the “Banque Nationale des ventes de produits phytopharmaceutiques par les Distributeurs agréés (BNV-D Traçabilité - https://ventes-produits-phytopharmaceutiques.eaufrance.fr)” as released to the public by official agencies at the township level (i.e. according to postal code, https://www.data.gouv.fr/fr). These data were extracted for 2017 and 2018 (the year before and the year when sampling was conducted). The sales of GLY and GLUF were associated with each trapping location on the basis of township positioning. The sales of GLY were highly correlated between years at the township level (R2=0.950), rendering it impossible to study the influence of potential use intensity separately in the year before or in the year of sampling. The marketing authorisation of pesticides containing GLUF was withdrawn in France by October 2017. The quantities sold in 2017 were very low (i.e. 0.75 kg) and sporadic (i.e. only one quantity reported as sold in the townships studied) whereas in 2018, no sales of GLUF were reported in the townships of the study area of concern. The quantities sold in 2017 and 2018 were thus summed for each a.s. and classified into two categories. For GLUF, the categories were “0” and “0.75”. For GLY, the median of the summed quantities in the dataset was calculated at 7300, and the categories were set at “lower than 7000” ([4138-7000[, 2 townships) and “higher than 7000” ([7000-9366[, 3 townhips).
Chemical analyses of glyphosate, AMPA and glufosinate
Full detailed information about chemicals and reagents, sample preparation, methodological approaches used for the chemical analyses and issues related to metrology are provided in Sup Info Annex 1. Text A2.
Briefly, the surface of hair samples were decontaminated with sodium dodecyl sulfate solution and methanol (Duca et al., 2014) After decontamination, the samples were spiked with internal standard solution, and water was added before overnight extraction. The supernatants were evaporated to dryness, and borate buffer and FMOC-Chloride were added before derivatization and injection into the LC-MS/MS Waters system Acquity UPLC H-class system equipped with a Waters Acquity BEH C18 column (100 × 2.1 mm, 1.7 μm particle size) coupled to a Waters Xevo TQ-S triple quadrupole mass spectrometer. With each analytical run, quality controls consisting in hair samples supplemented at several concentrations (0.1 to 100 pg/mg) and a blank sample were included.
The concentrations are expressed as pg/mg hair. The limits of quantification (LOQ, established as the lowest concentration level presenting variability and accuracy < 25%.) and lowest detected values (Minimum) are presented in Table 1. We did not perform specific experiments and calculations to compute limits of detection (see Sup Info Annex 1. Text A2). Importantly, only one concentration value was reported below the lowest concentration in quality controls (the minimum GLY concentration, see Table 1). All values above the lowest analytical detected value were included as “detects” contrary to “nondetects”. All reported concentration values, including values between the LOQ and the lowest detected value, were included as continuous data. The use of concentrations below the LOQ has been shown to be a suitable method that may be preferred to the absence of data or other established approaches such as imputation-based methods (i.e. efficiency in terms of bias and precision) (Keizer et al., 2015).
Dose reconstruction to assess the risk of deleterious effects in small mammals
Given the concentrations of GLY, AMPA and GLUF in the keratinized tissues of small mammals, our study highlights the exposure of these animals. However, the toxicological meaning of such hair concentrations is hard to interpret with respect to toxicity benchmarks (which are expressed as daily doses in mg active substance per kg body weight) because relationships between residues in hair and exposure doses or between residues in hair and in those other organs are not known. To overcome this gap and assess whether exposure levels could reach toxic levels, approaches of reverse dosimetry have been applied to estimate levels of exposure in terms of daily dose intake, to obtain data comparable to toxicological thresholds. Different types of data (i.e. concentrations in mammal body fluids and hair, residues in viscera and body burden in small mammals, toxicokinetics in small mammal blood, pharmacokinetics of fast-elimination chemicals) that may be used for such purposes have been collected from the literature (Sup Info Annex 1. Text A3 for full description), leading to different approaches of back-calculations computed under several assumptions when there was a lack of available data. Five different approaches could be computed for GLY and one for GLUF (Tables 2 and 3).
The objective of our risk assessment was to determine the overall orders of magnitude of daily dose intake relative to the concentrations measured in hair to evaluate whether wild small mammals may experience deleterious toxicological effects. The lack of data and assumptions that had to be applied hamper the accuracy of the calculated dose values. The interpretation of the findings was thus developed to consider ranges and global patterns rather than refined (but biased) toxicological assessments. The doses calculated were compared with toxicity data using the usual and regulatory procedure of toxicity-exposure ratio (TER) calculation, with associated triggers of 5 for chronic toxicity data and of 10 for acute toxicity data. The doses were reconstructed for each species, using four levels of residue concentrations measured in hair: minimum, median, 3rd quartile and maximum values (only one value available in house mice for GLY and in house mice and shrews for GLUF). The dose of GLY was compared to five toxicological thresholds: the short-term dietary no observed effect level (short-term dietary NOEL), the chronic 21-day no observed adverse effect level (chronic NOAEL, which is the same value as the reproductive NOAEL), the long-term NOAEL, the no mortality dose to the Oregon vole, and a “chronic low dose” limit. Details about the values of the thresholds are provided in the Sup Info Annex 1. Text A3. The dose of GLUF was compared to four available toxicological thresholds: the acute oral median lethal dose, the short-term dietary NOEL, the chronic 21-day NOAEL, and the long-term NOAEL.
The process ended with the TER values computed for each approach, under different scenarios of assumption, for the different levels of residue concentrations measured in hair, for each species, for the different toxicological thresholds (see Tables 2 and 3 and details in Sup Info Annex 2). Finally, the number of cases where TER exceeded the triggers, which highlights the risk for toxic effects in small mammals, and the number of individuals involved were summarized.
Data analysis and statistics
Analyses were performed using R (3.3.1), with the additional packages “AICcmodavg”, “effects”, “EnvStats”, “forestmodel”, “ggeffects”, “ggplot2”, “gtsummary”, “multcomp”, “pgirmess”, and “questionr”. Since more than two individuals were obtained from the same trapline in only eight out of the 29 where the animals were captured, pseudoreplication was not considered an issue. Moreover, the traplines extended over 50 metres, which should limit the influence of spatial autocorrelation between individuals.
The statistical approaches were designed to handle left-censored data within a situation where the smallest observations of data are nondetects, with high proportions of nondetects, skewed distributions, and small sample sizes, which hampered the use of methods considered “gold standards” to manage left-censored data (ITRC, 2013; Shoari & Dubé, 2018). Following recommendations to analyse datasets including nondetected and left-censored observations, we (1) used tests dedicated to censored data when possible, (2) computed descriptive statistics separately on detection frequencies and on detected concentrations (for which median, interquartile range, and outer percentiles were reported), (3) performed group comparisons separately on detection frequencies and on detected concentrations, and (4) preferred nonparametric tests to analyse the combined dataset, including uncensored data and nondetects with the latter being substituted by 0 (ITRC, 2013; Shoari & Dubé, 2018). Compared with other methods (i.e. imputation- and nonimputation-based approaches), the traditional substitution approach has been shown to be appropriate for handling nondetects in a study that investigated a dataset of pesticide concentrations in pollen and nectar to assess the exposure of bumble bees (Adjei & Stevens, 2022). Substitution with a half limit of detection outperformed the other approaches in the study of Adjei and Stevens (2022), which used parametric tests (nested ANOVA). However, we preferred to use 0 because (1) we used all reported values, including those between the LOQ and lowest detected values, instead of data left-censored by the limit of detection (Keizer et al., 2015), (2) the lowest detected values reached low values that would have brought the value of half of the LOD close to 0, (3) this approach allows easy identifying nondetects in the databases and graphics, and (4) nonparametric tests were used to compute statistics on both nondetects and quantified values.
Comparisons between the distributions of the compounds were performed using Tarone-Ware two-sample test for censored data (i.e. GLY vs. AMPA, GLY vs. GLUF, and AMPA vs. GLUF), with nondetects set to the corresponding lowest detected value of the corresponding compound as needed in such tests, for all species together (the sample sizes were too small to run the tests by species). Correlations between raw values with nondetects set to 0 of the concentrations of the three compounds were investigated using Spearman’s rank correlation tests. The relationships between the concentrations of each compound were further studied according to species via generalized linear models (Gaussian family) on log-transformed data (log(x+1)) to meet the assumptions of normality and variance homogeneity. The models were checked for homogeneity of variance, normality of error and linearity/additivity and leverage was checked (i.e. standardized residuals vs. leverage). An extreme outlier of GLY concentration was identified (one Microtus vole) as having overinfluence on the estimated parameters (i.e. Cook distance greater than 1). This outlier was removed from the dataset to compute model outputs and graphics in this part of the analyses.
Then, statistical analyses were performed on (1) GLY, AMPA or GLUF frequencies of detection and (2) GLY, AMPA or GLUF concentrations quantified above the lowest detected value (i.e. in samples where the compound was detected, without nondetects). According to the preliminary analyses showing differences between species, further analyses were conducted taking species into account in statistics using the factor “species” first in the models with the other factors. Full models including all explanatory factors and first-order interactions could not be used due the large number of NAs. Preliminary tests performed on combinations of factors, including interactions, revealed that none of the interactions were statistically significant and that models without interactions better fit the data (according to corrected Akaike information criterion (AICc) comparisons). The final models used to compute the statistics were therefore run without interactions.
We tested whether the detection of GLY, AMPA or GLUF in hair of small mammals was dependent on species, habitat, farming practices and proxies for treatment intensity using binomial GLMs (link = logit). The detection was coded as “0” for nondetected and “1” when detected. The models were built using species alone and species with each of the other variables as explanatory factors. The significance of the factors was checked by the chi-squared test. Odds ratios with confidence intervals were computed to characterize differences between levels.
We tested whether the concentrations of GLY, AMPA or GLUF in the hair of small mammals were dependent on species, habitat, farming practices and proxies for treatment intensity using GLMs on log-transformed values above the lowest detected value (i.e. without nondetects) to meet the assumptions of normality and variance homogeneity. The models were built using species alone and species with each of the other variables as explanatory factors. The significance of the explanatory factors was checked via an F test, and statistical differences between levels were via a t test.
To support the results and interpretation, both the detection and concentration data were analyzed via nonparametric tests on all of the data, with nondetects included and set to 0. The differences between species, habitats, farming practices, and the proxies of treatment intensity were tested adequately via the Wilcoxon-Mann-Whitney test or the Kruskal-Wallis test adequately depending on the number of levels of the factors (i.e. two or more). Post hoc multiple comparison tests after Kruskal-Wallis tests were used to investigate pairwise comparisons when relevant. Correlations between concentrations and landscape OF proportions or sales of a.s. used as raw continuous data were tested via Spearman's rank correlation test.
Results & Discussion
Detection frequency of GLY, AMPA and GLUF in the hair of small mammals
Considering all of the species together, GLY, AMPA and GLUF were detected in 64%, 51%, and 44% of the individuals, respectively (Table 1).
Compared with the quantities sold (in kg) in the townships studied, the quantities of GLUF were 2484 to 4844 times lower than those of GLY in 2018 and 1653 to 4548 times lower in 2017. Indeed, after its ban in 2017, GLUF could only have been used until stock depletion or usage derogation. Focusing on the part of the region where the study was carried out (i.e. “Département des Deux-Sèvres”), the purchases of GLUF rapidly decreased after 2015 and reached 0 in 2019 (BNV-D Traçabilité). Despite its low intensity of application compared with that of GLY, GLUF was detected in approximately half of the small mammal individuals.
Within the “PING” research programme (see M&M), the part of the study conducted over the same year over the same sampling sites as the present one detected GLY in 88% of the soil samples and 74% of the earthworm samples, AMPA in 58% of the soil samples and 38% of the earthworms, and GLUF in 35% of the soil samples and 12% of the earthworms (Pelosi et al., 2022). These detections in soil, earthworms and small mammals collected from arable fields and hedgerows, both in conventional and organic farming fields, revealed a generalized occurrence of GLY, AMPA and GLUF in the agricultural environment and fauna across this typical French agricultural area. In comparison, in three county-level hair pools of carcasses of big brown bats (Eptesicus fuscus; an insectivorous species weighing 15 to 26 g) in Missouri (USA), GLY was detected in all three analysed sample pools (100% detection frequency) (Hooper et al., 2022). The frequency of exposure to GLY in a farmland herbivorous mammal, the Iberian hare (Lepus granatensis) via analyses of residues in samples of gastric content, reached 45% of individuals found dead in March-April due to roadkill or unknown causes (Martinez-Haro et al., 2022), which is comparable to our results. In hunted hares the frequency of detection in gastric content ranged between 9% and 22% in animals from pesticide-treated areas. These later values in hunted hares look lower than the frequencies found in rodents in our study. This, possibly because residues in gastric content represent a snapshot of recent exposure whereas measurements in hair represent a temporally integrative exposure assessment during the time of hair growth, and because of the seasonal difference. Indeed, the hunted Iberian hares were collected from October to January, while our sampling session was conducted during springtime. The authors indicated that the frequency of detection in hares progressively increased over the hunting season, being 0% and 11% in October and November and 50% in December and January (Martinez-Haro et al., 2022). Issues related to local contamination may further explain the lower detection frequency in hunted hares.
Concentrations of GLY, AMPA and GLUF in the hair of small mammals
With respect to residue concentrations above the lowest detected value, we found that the three compounds presented overall similar ranges of concentrations, and the highest concentrations were reached for GLY, with values varying from 0.018 to 522 pg/mg (median = 2.65). The concentrations of AMPA ranged from 0.240 to 33.6 pg/mg (median = 1.39), and those of GLUF ranged from 1.16 to 25.5 pg/mg (median = 3.51) (Table 1). The concentrations of GLY did not differ significantly from those of AMPA or GLUF, and the concentrations of AMPA did not differ from those of GLUF (Tarone-Ware tests, p-values > 0.05, Sup Info Annex 1. Text A4).
In county-level hair pools from big brown bats in Missouri, Hooper et al. (2022) reported concentrations of GLY varying from 5.7 to 4505.2 pg/mg. These maximum concentrations are much higher than those measured here in shrews (median = 0.8 pg/mg) and almost ten times higher than the maximum concentration measured in our study in a vole (i.e. 522 pg/mg). Several nonexclusive hypotheses may be proposed to explain such high levels found in bats: analytical issues, physiological and ecological traits of the species enhancing exposure and/or bioaccumulation (Fritsch et al., 2024a), and issues related to the amount of GLY used and the timing of exposure of the animals after GLY application.
Newton et al. (1984) studied the fate and behaviour of GLY and AMPA in forest brush field ecosystems in the Oregon Coast Range (USA) following aerial application of GLY at doses comparable to the recommended GLY application rate in the EU (i.e. 3.3 kg a.s./ha there, maximum field application of 4.32 kg a.s./ha in the EU (European Commission, 2015)). They measured the residues of GLY and AMPA in the viscera and body of free-ranging small mammals captured at the sites before treatment and over the 55 days posttreatment. As in the present work, GLY was detected in all of the trophic groups (LOD of 0.10 mg/kg = 100 pg/mg). However, AMPA was detected in two voles only at 130 and <160 pg/mg (LOD of 100 pg/mg) (Newton et al., 1984), which is far lower than in our study. This may be because the lowest detected value in our work was much lower (i.e. 0.240 pg/mg) and because different tissues were used to measure residues.
Interestingly, considering these previous findings of Newton et al. (1984) in forest small mammal tissues, the concentrations varied over the same order of magnitude as the maximum concentrations measured in our hair samples from agricultural landscapes. They reported concentrations in viscera ranging from < 100 pg/mg (LOD) to 1690 pg/mg in shrews, 5080 pg/mg in deermice and 1700 pg/mg in voles. They reported concentrations in body tissues ranging from < 100 to 410 pg/mg in shrews, 400 pg/mg in deermice and 250 pg/mg in voles. As a comparison, Martinez-Haro et al. (2022) reported GLY concentrations ranging from 0.11 to 16 μg/g (110 to 16000 pg/mg) in the gastric content of Iberian hares found dead or hunted in Spain. Newton et al. (1984) showed that the concentrations were the highest during the first days following herbicide treatment both in the viscera and body tissues. Body concentrations drecreased under the LOD by 2 weeks after treatment while they were still detectable in the viscera, and concentrations in the viscera decreased to nondetectable concentrations or near the LOD by day 55 after treatment (Newton et al., 1984). The environmental persistence of GLY is estimated to vary around 43 days (reported values ranged from 1 to 180 days, up to 197 days) (Gandhi et al., 2021; Kissane & Shephard 2017). Although residues may be detected several months after application following spraying and rainfall, the binding to soil and biodegradation of glyphosate is estimated between 4 to 11 days post-treatment (Kissane & Shephard 2017). In their forest study, Newton et al. (1984) reported GLY half-lives ranging from 10 to 27 days in foliage and litter, half-life of 40 days for exposed soil, and half-life of 29 days for litter-covered soil. The GLY residues overall dropped under 20-30% of the initial deposit within 7-14 days (Newton et al., 1984). This pattern of GLY concentration decrease over time in the environment and in small mammal body and viscera following a single GLY application, together with differences in the matrices analysed, may explain the overall lower GLY levels in our study in comparison to the one of Newton et al. (1984).
This also suggests that the highest concentrations found here may correspond to animals highly exposed to GLY soon after its application when environmental concentrations were the highest, and/or repeatdly exposed during the growth of hair. Our samples were collected during springtime, from May-June, whereas glyphosate is usually applied to winter cereals during late summer or fall (often in October). The seasonal moult of small mammals is expected to be completed mostly by May-June, when they were captured, and the sampled fur might have partly or even totally grown within the period of February-May (Sealander, 1972; López-Fuster et al., 1986; Abad, 1991). However, the postwinter moult may be incomplete, depending on the age of individuals and the species of concern, allowing the possibility for hair grown during autumn to be included in the analysed fur sample (Beltran et al., 2018).
Correlations between the concentrations of GLY, AMPA and GLUF
The concentrations of the three compounds in the hair of small mammals were significantly related (Fig. 2, Annex Fig. A1). Positive correlations were observed between AMPA and GLY (correlation coefficient Spearman’s rho=0.84), between GLUF and GLY (rho=0.79), and between AMPA and GLUF (rho=0.89) (Annex Fig. A1). The variance explained (cf. determination coefficient R2) was greater for the factor species (0.41 to 0.42) than for compounds (0.16 to 0.27) (Fig. 2).
The correlation between AMPA and GLUF differed only according to species, with greater slopes for shrews and voles than for the two species of wild mice (Fig. 2). The correlations between AMPA and GLY and between GLUF and GLY were similar between species (Fig. 2).
The strength of the association between AMPA and GLUF was greater than that between AMPA and GLY (i.e. greater rho values, greater R2 values; Fig. 2, Sup Info Annex 1. Fig. A1), and the relationship presented greater slope coefficients. This may be seen as unexpected since AMPA is a GLY transformation product.
Such patterns of correlation between the compounds may reflect different exposure pathway contributions between GLY versus AMPA and GLUF. As GLY is poorly metabolized in mammals (EFSA, 2015a), the origin of AMPA should be external (environmental) rather than internal (degradation of GLY in the body), as suggested in humans (Alvarez et al.,2022). Studying GLY and AMPA residues in human hair, Alvarez et al. (2022) reported different origins of GLY and AMPA because AMPA was rarely detected in comparison with GLY in both farmers and nonoccupationally exposed subjects, and the ratio of GLY to AMPA greatly differed between the subjects. The correlation between GLY and AMPA or between GLY and GLUF likely originates from a spatial relationship related to the geographic occurrence of the herbicidal treatments, which increases the probability of co-occurrence of parent and transformation products in herbicide-treated areas. As hypothesized, small mammals may be exposed to GLY via the dietary route but also via direct spraying if they are present in the plots during the treatments and orally after spraying treatments through grooming activities. Conversely, dietary exposure, related to the accumulation of the compounds in the soil, biota and food webs, may be the predominant exposure pathway to AMPA and to GLUF (whose use as a PPP is no longer approved) in small mammals.

Figure 2 - Correlations between the concentrations of glyphosate, AMPA and glufosinate in the hair of small mammals. The concentrations are expressed in pg/mg. The statistical outputs of general linear models are provided on graphs. Predictions are plotted according to species when the interaction between the factors is significant, and average predictions are provided when the interaction is not significant.
Differences between species in detection frequency and concentration of the compounds
The frequency of detection differed among species for the three compounds (Fig. 3, Sup Info Annex 1. Table A2). In all cases, the odds ratios were significantly different between the voles and wood mice (Sup Info Annex 1. Table A2). The probabilities of detection of GLY, AMPA and GLUF were significantly greater in common and bank voles than in shrews and wood mice (Fig. 3). The results were contrasted for the house mouse, likely because of the small sample size (Fig. 3).
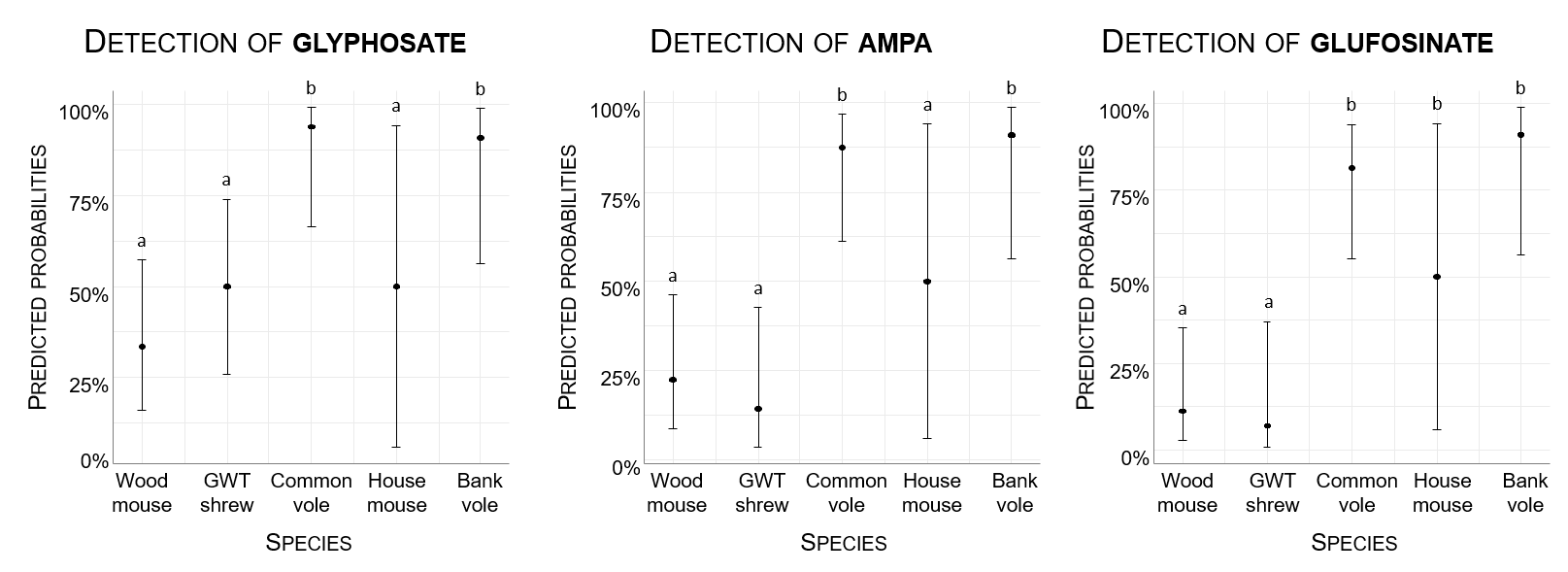
Figure 3 - Probabilities of the detection of glyphosate, AMPA and glufosinate in the hair of small mammals according to species. The predicted probabilities of detection were obtained from the statistical binomial models ’’Detection ~ Species’’. Statistical outputs of comparisons between levels are provided, and the species sharing similar letters did not differ statistically. Abbreviations: GWT shrew stands for greater white-toothed shrew. The sample sizes are provided in Table 1.
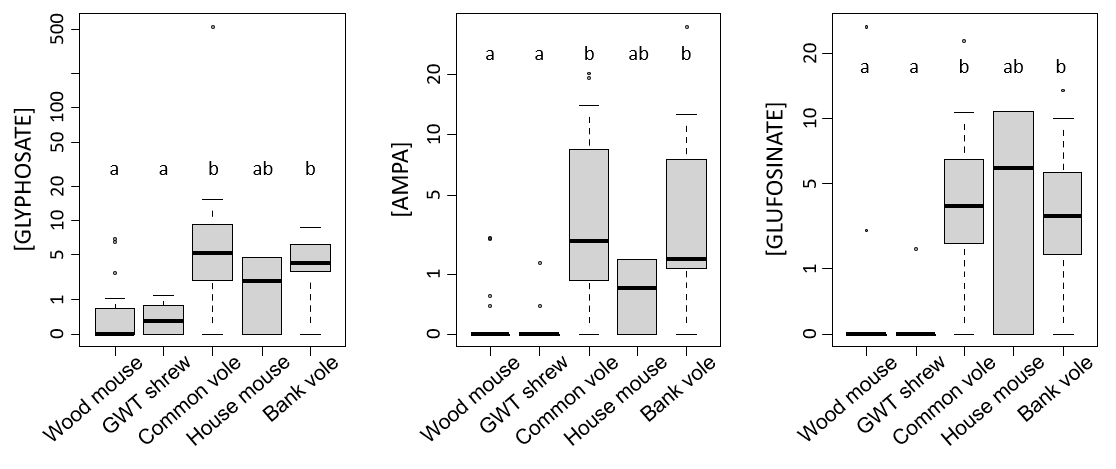
Figure 4 - Concentrations of glyphosate, AMPA, and glufosinate in the hair of small mammals according to species. The concentrations are expressed in pg/mg. Nondetects are set to 0. Statistical significance is indicated with letters, and different letters between levels indicate significant differences. Statistics were perfomed via the Kruskal-Wallis test followed by posthoc multiple comparison tests to investigate pairwise differences. Abbreviations: GWT shrew stands for greater white-toothed shrew. The sample sizes are provided in Table 1.
When analyses were run on the subset of data with quantified concentrations in small mammals (i.e. without nondetects), the concentrations of GLY, AMPA and GLUF did not significantly differ among species (Sup Info Annex 1. Table A3). However, the trends considering GLY and AMPA were comparable overall, since estimates for concentrations were found to be the lowest in shrews and wood mice for GLY and the highest in voles for AMPA (Sup Info Annex 1. Table A3). Considering GLUF, the highest estimates were observed for wood mice and house mice (Sup Info Annex 1. Table A3). When the tests included nondetects set to 0, the results revealed higher levels of GLY, AMPA, and GLUF in common voles and bank voles than in wood mice and shrews (Fig. 4). The concentrations in house mice did not significantly differ from those in other species, with intermediate values (note that the sample size is small) (Fig. 4).
In their forest study where residues of GLY and AMPA were measured in several free-ranging mammalian herbivores, carnivores, and omnivores, Newton et al. (1984) reported that GLY was present in all trophic groups, whereas AMPA was present in only a few herbivorous voles. Our results are in line with these findings, with confirmed exposure to GLY (and AMPA) in both herbivorous and granivorous or omnivorous rodents and insectivorous shrews and greater levels of AMPA in herbivorous mammals (note that the bank vole was often considered a granivore, but recent literature has shown omnivory (Balčiauskas et al., 2022)).
Newton et al. (1984) emphasized that exposure to and accumulation of GLY in mammals varied according to diet preference, since omnivores (deermice) had the highest concentrations of GLY in the viscera during the two weeks posttreatment, suggesting a higher initial intake of GLY in omnivores, but such visceral levels decreased the fastest in comparison to carnivores (shrews and weasel) and herbivores (squirrel, vole, chipmunk). The detailed data revealed greater body concentrations in shrews and deermice (0.35 to 0.41 mg/kg) than in squirrels and voles (0.13 to 0.25 mg/kg) within the first days posttreatment. Our results similarly revealed differences in the accumulation or uptake of GLY and AMPA between trophic groups or species, but various trends were identified according to trophic preferences. Overall, we found lower detection frequencies and concentrations in insectivores, whereas the highest detection frequencies and concentrations were observed in herbivorous and omnivorous voles. Both the detection probability and the concentrations were greater for the two species of voles than for the omnivorous wood mice and house mice in our study. The differences in the matrices analysed and in the timing of exposure after application may explain the discrepancies between the findings of Newton et al. (1984) and ours regarding the influence of diet on accumulation. Moreover, differences in the identities of the species (i.e. European vs. American species) and ecosystems (i.e. agrosystem vs. forest) may explain the discrepancies.
The physiological and metabolic traits affecting uptake and accumulation in body tissues/fluids and the ecological traits affecting exposure may shape the differences found here between species or trophic groups, but their role is difficult to disentangle. The species exhibiting the lowest detection frequency differ in terms of phylogeny, since both Apodemus and Mus mice of the subfamily Murinae as well as Crocidurinae Crocidura shrews were represented (Sup Info Annex 1. Fig. A4). They also differ in their ecological traits since they are omnivorous and insectivorous. The most herbivorous and, to a lesser extent, the least mobile species, which are the two species of voles studied here, presented the highest detection frequencies and greatest concentrations. Microtus and Myodes voles are classified into two different genera, but both belong to the same subfamily, Arvicolinae (Sup Info Annex 1. Fig. A2). The correlation between the levels of GLUF and AMPA varied under species-specific patterns, as detailed above. The wood mouse and the house mouse, which are both omnivorous Murinae species, presented a slope different from that of the herbivorous common vole, the omnivorous bank vole and the insectivorous greater white-toothed shrew which presented a steeper slope. The bank vole is usually classified as showing intermediate features of traits between the wood mouse and the common vole in terms of mobility and homerange size, habitat selectivity, and diet breadth (van den Brink et al., 2011). According to ecotoxicological studies where the dietary route is the predominant exposure pathway, its responses are expected to be intermediate or closer to the trends exhibited by the wood mouse than by the common vole (van den Brink et al., 2011; Baudrot et al., 2018).
Although the importance of toxicokinetics on accumulation in hair cannot be ignored (Faÿs et al., 2023), our results may again suggest the involvement of different origins of the compounds and of a different contribution of the various exposure routes in shaping interspecies accumulation dissimilarities. The differences found here between species and between animals captured in different habitats within the landscape mosaic highlights the need to include both ecological and physiological features in the wildlife risk assessment process and postregistration surveys of the unintentional effects of PPP (Morrissey et al., 2023). Importantly, these results highlight the need for additional research to gain knowledge about the traits that shape the exposure and uptake of PPP in wildlife and about all the interacting mechanisms involved.
Influence of habitat, farming practices, and proxies of treatment intensity on the detection frequency of the compounds
The frequency of detection did not differ according to habitat or farming practices (Sup Info Annex 1. Table A2). Accordingly, the study of Pelosi et al. (2022), which was done within the same research project as the present study (same location of sampled plots and sampling in 2018) did not show differences in GLY detection frequency in soils sampled from cereal fields or hedgerows. However, in earthworms, GLY and AMPA were detected more frequently in cereal fields than in hedgerows.
The frequency of detection of GLY, AMPA and GLUF in small mammals was not significantly dependent on the proxies of treatment intensity at the plot, landscape and township scales, except in the case of GLUF sales (Sup Info Annex 1. Table A2, Fig. A3, Fig. A4). However, such an effect of a.s. sales on GLUF detection probabilities was negative, i.e. the detection probabilities were lower in townships where GLUF sales were the highest, and the differences between categories were not significant (Sup Info Annex 1. Table A2, Fig. A4). To investigate this paradoxical result further and avoid any interpretation bias, a model with GLUF sales as the only explanatory factor (i.e. without the factor “species” first in the model formula) was run, revealing an absence of significant differences (Sup Info Annex 1. Table A2, Fig. A4).
Hedgerows, woodlots and fields cultivated under OF are not supposed to be targeted by pesticide treatments, but they may be unintentionally contaminated due to drift or run-off (Gandhi et al., 2021). Furthermore, the home range of small mammals can overlap wooded patches, nontarget crops and surrounding cropped habitats. In agricultural topsoil samples from conventional and organic farms across Europe screened for chemicals, GLY and AMPA were among the compounds with the highest frequency of detection and the greatest concentrations (Geissen et al., 2021). Moreover, AMPA was the residue most often detected in both conventional and organic fields, with 96% occurrence in conventional systems and 83% in organic systems (Geissen et al., 2021). AMPA and GLY were also the most commonly detected pesticides in soils under various land uses sampled over mainland France, in which 111 pesticide residues were screened (Froger et al., 2023). The detection frequency was 83% for AMPA and 70% for GLY, and the two compounds were found both in treated cultivated fields and nontargeted habitats, as they occurred both in conventional and organic farming plots, and in different types of land uses: arable lands, orchards, forests, grasslands, and brownfields (Froger et al., 2023). Such a global occurrence of these two compounds, including in organic fields and grasslands or forests, echoes the pervasive exposure found here in small mammals.
Martinez-Haro et al. (2022) reported results contrasted with the present findings about the influence of farming practices on exposure to GLY: the prevalence was 9–22% in the gastric content of hunted Iberian hares and 45% in hares found dead in pesticide-treated areas, whereas no residues were detected in hares from organic crops (pesticide-free areas). As stated above, this pattern may be due to the differences in the matrices analysed which reflect different temporal integration of exposure. Finally, the intensity and extent of the GLY treatments may have been greater in our study region, and the OF and CF plots may have been geographically closer in our case.
In addition to the study of GLY levels in environmental matrices such as soil or biota, more research is needed to characterize the patterns of exposure of wildlife to GLY, AMPA and GLUF and quantify the underlying processes and spatiotemporal dynamics.
Influence of habitat, farming practices, and proxies of treatment intensity on the concentrations of the compounds
In terms of the concentrations of the compounds when quantified, the GLY concentrations differed among the small mammals captured in the different habitats: they were greater in individuals from hedgerows than in those from cereal fields (Fig. 5, Sup Info Annex 1. Table A3, Fig. A3). Considering both nondetects set to 0 and quantified concentrations, the trends were similar but not significant (Fig. 6). Such a pattern was not observed in the cases of AMPA and GLUF (Fig. 6, Sup Info Annex 1. Table A3).
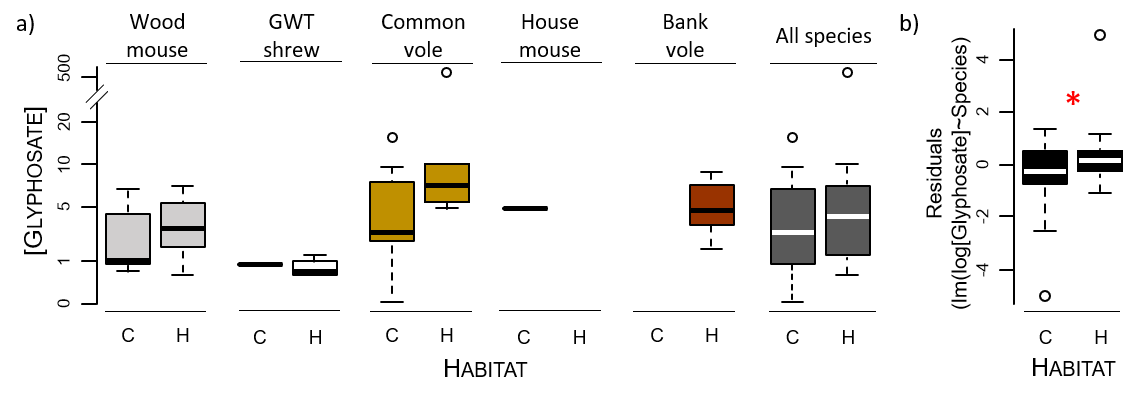
Figure 5 - Concentrations of glyphosate in the hair of small mammals without nondetects (a) according to species and habitat and (b) according to habitat taking species into account statistically in modelling. The concentrations are expressed in pg/mg in hair. The statistical significance is depicted by a red asterisk. The habitats are represented as “C” for cereals and “H” for hedgerows. The GWT shrew stands for the greater white-toothed shrew. The sample sizes are provided in Table 1 and Table A1.
This may be a confirmation of the different contributions of exposure pathways for GLY in comparison to AMPA and GLUF. This pattern may indeed be related to a contribution of overspray in addition to dietary exposure because of direct spraying and/or elevated GLY deposits on hedgerow vegetation (1) during spraying due to drift and/or (2) due to (wet) deposition (Gandhi et al., 2021). GLY is known to have a low volatility but is found in air and rainfall in intensively treated areas, at relatively high frequencies and concentrations, and is subjected to wet deposition (Chang et al., 2011). Compared with those in open habitats, contaminant deposits, including PPP, on vegetation in hedgerows are known to be enhanced by larger surfaces of interception and by “edge effect” (Davis et al., 1994; Ould-Dada et al., 2002; Fowler et al., 2004; Lazzaro et al., 2008). Mayer et al. (2020) reported for European hares (Lepus europaeus) that pesticide uptake by foraging could be 7-fold lower than uptake via overspray/oral grooming. Moreover, GLY has been found to persist in vegetation for up to one year following treatments in perennial forested areas, representing a potent dietary source of exposure for wildlife (Edge et al., 2021).
In a previous study addressing the exposure of small mammals to banned and currently used PPP by residue screening in hair, which was based on animals sampled from the same study area and another region in France, we reported greater contamination (in terms of the number of compounds and concentrations) in individuals captured in hedgerows than in those captured in grasslands. However, the contamination did not differ between animals captured in hedgerows and those sampled from cereal crops (Fritsch et al., 2022). In Pelosi et al. (2022), GLY and AMPA concentrations in both soils and earthworms were, however, higher in samples from cereal fields than in those from grasslands and hedgerows. This may be an argument in favour of the contribution of processes related to overspraying in determining small mammal exposure to GLY. The role of external exposure to herbicides in small mammals and the importance of oral exposure by grooming remain to be investigated. The contribution of nondietary routes to wildlife exposure is still an overlooked field of research that needs to be addressed.
No differences in the quantified concentrations of GLY, AMPA or GLUF in the hair of small mammals were detected according to farming practices or proxies of pesticide treatment intensity at the different scales (Sup Info Annex 1. Table A3). Running the analyses on the basis of concentrations and including nondetects as 0 provided similar results (Fig. 6, Sup Info Annex 1. Fig. A3).
Previously, we also highlighted the absence of differences in contamination by currently used pesticides other than GLY between small mammals captured in conventional or organic farming plots (neither the number of compounds nor their concentrations in individuals) (Fritsch et al., 2022). In soils and earthworms sampled within the ZAPVS, Pelosi et al. (2022) also reported no significant differences in GLY, AMPA or GLUF levels according to pesticide use (i.e. comparing treated conventional fields to nontreated habitats such as grasslands, hedgerows and organic fields) or according to farming systems in cropped fields (i.e. fields cropped under conventional or organic farming).
The ubiquity of exposure to GLY, AMPA and GLUF in arable landscapes, both in crops and agroecological infrastructure, was therefore shown in recent studies for soils and for both terrestrial invertebrates and vertebrates belonging to various taxa and of different trophic levels. The transfer of GLY, AMPA and GLUF in food webs is thus questioned and requires further attention. This widespread occurrence of GLY, AMPA and GLUF in the environment and biota, regardless of the farming practices and intensity of use at the landscape scale, indicates pervasive contamination that may reflect the magnitude of repeated GLY-based (and formerly GLUF-based) herbicidal use over large surfaces in French agrosystems.
On the basis of our results and recent literature, the beneficial role of agroecological infrastructures (e.g. hedgerows) and nontreated crops as refuges or recovery areas towards pesticide treatments in agroecosystems under the current agricultural model is questioned. Our findings highlight that the current surfaces of nontreated habitats within the agricultural landscapes studied are insufficient to mitigate the exposure of wildlife to GLY, AMPA and GLUF. We reached similar conclusions when we studied the exposure of small mammals to 73 residues of fungicides, herbicides (excluding GLY, AMPA and GLUF), insecticides, and some of their metabolites in Fritsch et al. (2022). With respect to the mitigation of unintentional effects of PPP on wildlife, these results call for a reassessment of the relevance and efficiency of the current procedures and a strengthening of national and regional initiatives for agri-environmental schemes.
Further investigations into the pathways of wildlife exposure to PPP and responses at the individual and population levels are needed to understand and predict the role of agroecological infrastructures in shaping the direct and indirect effects of PPP. The beneficial role of hedgerows in limiting inputs of PPP on non-target plots around treated fields can be leveraged in agri-environmental schemes, but this desirable use of agroecological infrastructures to protect the environment and biodiversity may turn seminatural habitats into “ecotoxicological traps”. These issues deserve attention and should be addressed in both future research studies and monitoring schemes.
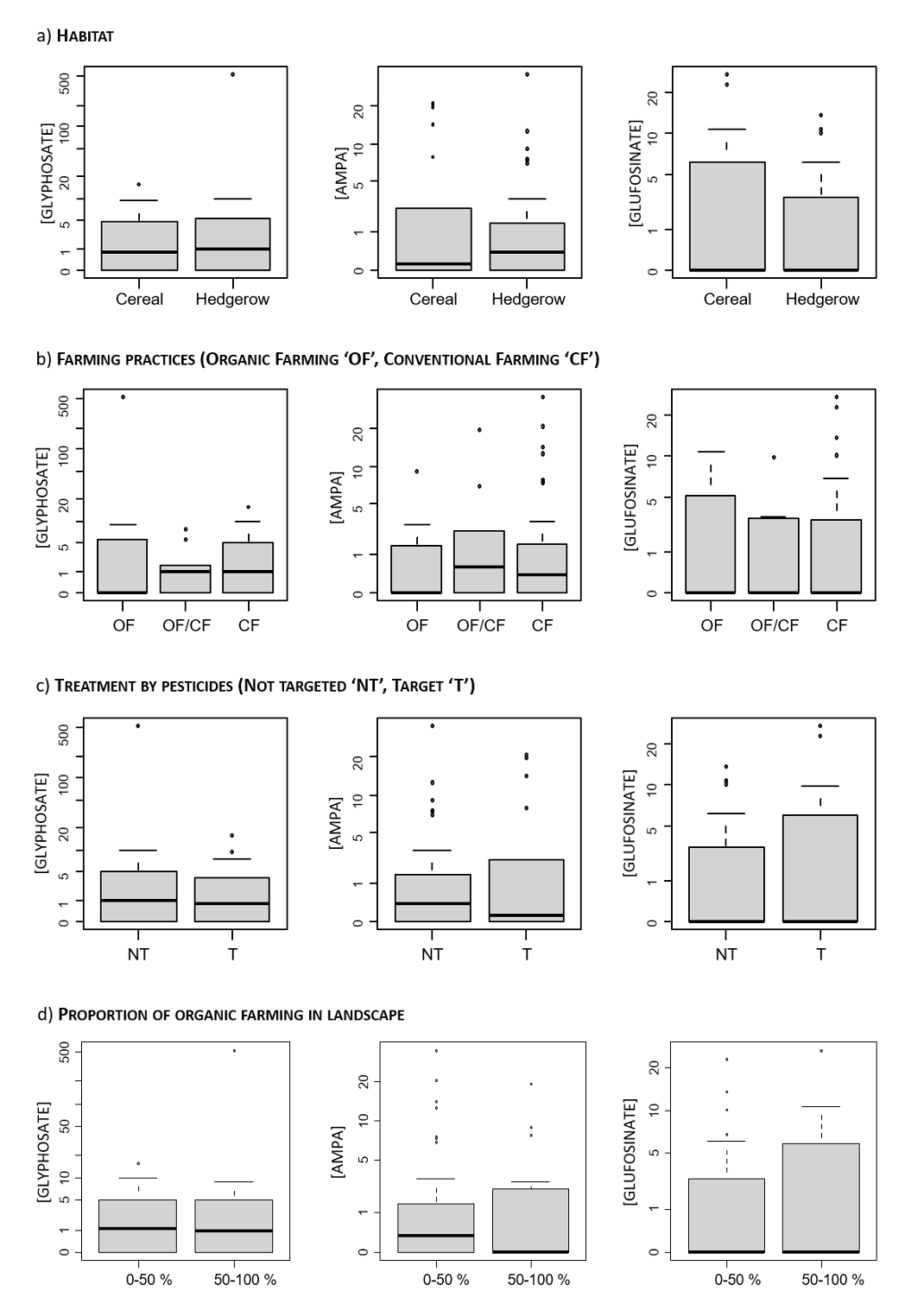
Figure 6 - Concentrations of glyphosate, AMPA, and glufosinate in hair of small mammals with nondetects set to 0, according to habitat where captured (a), farming practices where captured (b), and proxies of treatment intensity at the plot scale (c) and at the landscape scale (d). The concentrations are expressed in pg/mg. Statistics were perfomed via the Wilcoxon-Mann-Whitney test or the Kruskal-Wallis test depending on the number of levels of the factor of concern. Posthoc multiple comparison tests after Kruskal-Wallis tests were used to investigate pairwise comparisons when relevant. No significant differences were found.
Dose reconstruction for risk assessment
To evaluate the risk of unintentional impacts in small mammals by comparing exposure dose values estimated from back-calculation using the concentrations measured in hair to toxicological data, we found that toxic thresholds could be exceeded for both GLY and GLUF.
In the case of GLY, at least one of the five considered toxicological thresholds was exceeded according to the trigger of concern for at least one species in all of the approaches (Table 2). The potential for toxicological impairment was thus observed in all of the approaches developed. This finding suggests a risk for small mammals, especially rodents, and particularly for herbivorous voles (Table 2). Often, only the maximum value measured in Microtus voles reached toxic levels (10 times out of 16 scenarios). In five of the 16 scenarios computed via the different approaches, all of the species were found to be at risk, with maximum but also median and minimum concentrations measured being involved. At the population level, in the “best-case” scenario, toxicological effects related to GLY exposure impaired at least 6% of individuals in the common voles studied here, which represents a risk of effects in 2.1% of the rodents and in 1.6% of the small mammal community studied, including insectivorous species (Table 2). In the worst-case scenario, toxicological impairment occurred in 87.5% of the common voles, 88.9% of the voles (bank voles and common voles), 66% of all of the rodent species, 50% of the shrews and 62.3% of the individuals within the small mammal community (Table 2). Up to 93.8% of the common voles and 92.6% of the individuals of all vole species may be impacted if individuals close to the trigger value of 5 are included in the calculation (Table 2).
From GLY concentrations in gastric content, Martinez-Haro et al. (2022) calculated a daily dose for Iberian hares of 0.2-32.8 mg of GLY/kg body weight/day, with a median estimated at 0.36 mg GLY/kg body weight/day. We found here estimated daily doses varying from 0.00002 to 1231 mg GLY/kg body weight/day (Table 2). The median for all approaches was 0.373 mg GLY/kg body weight/day (1st quartile = 0.037 mg GLY/kg body weight/day, 3rd quartile = 1.84 mg GLY/kg body weight/day). One may notice the congruence between these estimated median values of exposure dose in Iberian hares and in small mammals here.
For GLUF, estimated daily doses varied from 0.080 to 6.38 mg GLUF/kg body weight/day (Table 3). The median for all approaches was 0.811 mg GLUF/kg body weight/day (1st quartile = 0.314 mg GLUF/kg body weight/day, 3rd quartile = 1.57 mg GLUF/kg body weight/day). The four toxicological thresholds considered were exceeded in the two scenarios for all rodent species, including the maximum concentrations measured in our samples but also the third quartile, median and even minimum values (Table 3). This would translate, on the basis of our captured individuals, into a possible impairment related to GLUF exposure in 31.3 to 75% of the common voles, 25.9 to 70.4% of the vole individuals, 19.1 to 46.8% of individuals within the rodent population and 14.8 to 36.1% of individuals within the small mammal community (Table 3). This result suggests that many of the free-ranging nontargeted small mammals could be at risk for toxicological effects related to exposure to GLUF, especially herbivorous and granivorous or omnivorous species, at the time of sampling (i.e. 2018), although the use of this compound in France has largely decreased since 2015.
Thus, our findings are consistent with the first conclusions of the risk assessments performed during the registration processes, which identified potential threats for both GLY and GLUF to herbivorous, granivorous and insectivorous terrestrial vertebrates, especially herbivorous voles (EFSA, 2005, 2012, 2015a). Note that the regulatory assessments were based on dietary exposure, but wildlife may be exposed to PPP - and this is especially the case for GLY when used through spraying - via additional routes such as oral uptake by grooming, inhalation, dermal contact, and/or ocular contact, as mentioned for human populations (Mineau, 2011; Mayer et al., 2020; Gandhi et al., 2021).
Table 2 - Summary of risk assessment for glyphosate based on reverse dosimetry approaches. Details are provided in Sup Info Annex 1 and Annex 2. Abbreviations: a.s. active substance; Min minimum; Max maximum; Med median; Nb number; Ind individuals; L-T NOAEL long-term no observed adverse effect level; S-T NOEL short-term dietary no observed effect level; Ac NOAEL acute NOAEL;Ch NOAEL chronic NOAEL; CLD chronic low dose threshold; Apsy Apodemus sylvaticus (wood mouse); Crru Crocidura russula (greater white-toothed shrew); Miar Microtus arvalis (common vole); Mumu Mus musculus (house mouse); Mygl Myodes glareolus (bank vole); Max maximum; Q3 3rd quartile; Med median; Min minimum; PK pharmacokinetics.
Approach Scenarios Type of data / Specific data /Assumptions | Estimated dose (mg a.s./kg/day) Min-max [med] | Cases when toxicity/exposure ratio exceed trigger value (of 10 for acute toxicity data or of 5 for chronic toxicity data) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Nb cases | Threshold of concern | Species of concern | Level [C]hair of concern | Nb ind | Nb rodents | Nb shrews | % All species | % Rodents | % Voles | % Microtus | % Shrews | ||
(including individuals close to the trigger*) | |||||||||||||
Approach 1 [GLY] in human urine and hair | |||||||||||||
Scenario 1.1 Head hair | |||||||||||||
Scenario 1.1.a Urinary excretion 10% | 0.0008 - 23.9 [0.186] | 3 | L-T NOAEL, S-T NOEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 4 | 6 | ||
Scenario 1.1.b Urinary excretion 20% | 0.0004 - 11.9 [0.093] | 2 | L-T NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 4 | 6 | ||
Scenario 1.1.c Urinary excretion 1% | 0.0082 - 239 [1.86] | 16 | L-T NOAEL, S-T NOEL, Ac NOAEL, Ch NOAEL, CLD | Miar, Apsy, Mumu, Mygl, Crru | Max, Q3, Med | 29 (33) | 25 (26) | 4 (7) | 48 (54) | 53 (55) | 74 (78) | 69 (75) | 29 (50) |
Scenario 1.2 Pubic hair | |||||||||||||
Scenario 1.2.a Urinary excretion 10% | 0.0042 - 123 [0.956] | 12 | L-T NOAEL, S-T NOEL, Ac NOAEL, Ch NOAEL, CLD | Miar, Apsy, Mygl | Max, Q3, Med | 16 (18) | 16 (17) | (1) | 26 (30) | 34 (36) | 48 (48) | 50 | (7) |
Scenario 1.2.b Urinary excretion 20% | 0.0021 – 62 [0.478] | 4 | L-T NOAEL, S-T NOEL, Ac NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Scenario 1.2.c Urinary excretion 1% | 0.0424 - 1230.84 [9.56] | 31 | L-T NOAEL, S-T NOEL, Ac NOAEL, Ch NOAEL, CLD | Miar, Apsy, Mumu, Mygl, Crru | Max, Q3, Med, Min | 37 | 30 | 7 | 61 | 79 | 85 | 81 | 50 |
Approach 2 Body burden in small mammals 7 days after dose | |||||||||||||
Scenario 2.1 Proportions hair/body burden | 0.0018 - 52.4 [0.366] | 4 | L-T NOAEL, S-T NOEL, Ac NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Scenario 2.2 [C]whole body = [C]hair | 0.0018 - 52.2 [0.365] | 4 | L-T NOAEL, S-T NOEL, Ac NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
*Values considered close to the trigger: less than 1 of difference, e.g. value of 5.2 for a trigger of 5
Table 2 - continued
Approach Scenarios Type of data / Specific data /Assumptions | Estimated dose (mg a.s./kg/day) Min-max [med] | Cases when toxicity/exposure ratio exceed trigger value (of 10 for acute toxicity data or of 5 for chronic toxicity data) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Nb cases | Threshold of concern | Species of concern | Level [C]hair of concern | Nb ind | Nb rodents | Nb shrews | % All species | % Rodents | % Voles | % Microtus | % Shrews | ||
(including individuals close to the trigger*) | |||||||||||||
Approach 3 Toxicokinetics in small mammal blood | |||||||||||||
Scenario 3.1 Anadon et al. (2009) | |||||||||||||
Scenario 3.1.a GLY residues | 0.0016 - 45.2 [0.316] | 4 | L-T NOAEL, S-T NOEL, Ac NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Scenario 3.1.b AMPA residues | 0.2307 - 32.3 [1.55] | 16 | L-T NOAEL, S-T NOEL, CLD | Miar, Apsy, Mumu, Mygl, Crru | Max, Q3, Med | 24 | 23 | 1 | 39 | 49 | 74 | 63 | 7 |
Scenario 3.2 Kim et al. (2023) GLY residues | <0.0001 - 8.54 [0.013] | 1 | CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Approach 4 Residues in viscera and body of small mammals | |||||||||||||
Scenario 4.1 Min ratio viscera/body residues | 0.0001 - 3.82 [0.0031] | 1 | CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Scenario 4.2 Average ratio viscera/body residues | <0.0001 - 8.54 [0.0069] | 1 | CLD | Miar | Max | 1 (2) | 1 (2) | 2 (3) | 2 (4) | (7) | 6 (13) | ||
Scenario 4.3 Max ratio viscera/body residues | 0.0004 - 11.4 [0.019] | 2 | L-T NOAEL, CLD | Miar | Max | 1 | 1 | 2 | 2 | 6 | |||
Approach 5 Prediction of incorporation in hair from PK | |||||||||||||
Scenario 5.1 Glyphosate residues | 0.921 – 523 [4.55] | 20 | L-T NOAEL, S-T NOEL, Ac NOAEL, Ch NOAEL, CLD | Miar, Apsy, Mumu, Mygl, Crru | Max, Q3, Med, Min | 38 (39) | 31 (32) | 7 | 62 (64) | 66 (68) | 89 (93) | 88 (94) | 50 |
Scenario 5.2 AMPA residues | 0.824 - 34.2 [1.95] | 20 | L-T NOAEL, S-T NOEL, CLD | Miar, Apsy, Mumu, Mygl, Crru | Max, Q3, Med, Min | 30 (31) | 28 (29) | 2 | 49 (51) | 60 (62) | 85 (89) | 81 (88) | 14 |
*Values considered close to the trigger: less than 1 of difference, e.g. value of 5.2 for a trigger of 5
Table 3 - Summary of risk assessment for glufosinate based on reverse dosimetry approaches. Details are provided in Sup Info Annex 1 and Annex 2. Abbreviations: a.s. active substance; Min minimum; Max maximum; Med median; Nb number; Ind individuals; L-T NOAEL long-term no observed adverse effect level; S-T NOEL short-term dietary no observed effect level; Ac NOAEL acute NOAEL;Ch NOAEL chronic NOAEL; CLD chronic low dose threshold; Apsy Apodemus sylvaticus (wood mouse); Crru Crocidura russula (greater white-toothed shrew); Miar Microtus arvalis (common vole); Mumu Mus musculus (house mouse); Mygl Myodes glareolus (bank vole); Max maximum; Q3 3rd quartile; Med median; Min minimum; PK pharmacokinetics.
Approach Scenarios Type of data / Specific data /Assumptions | Estimated dose (mg a.s./kg/day) Min-max [med] | Cases when toxicity/exposure ratio exceed trigger value (of 10 for acute toxicity data or of 5 for chronic toxicity data) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Nb cases | Threshold of concern | Species of concern | Level [C]hair of concern | Nb ind | Nb rodents | Nb shrews | % All species | % Rodents | % Voles | % Microtus | % Shrews | ||
(including individuals close to the trigger*) | |||||||||||||
Approach 1 Toxicokinetics in small mammal blood | |||||||||||||
Scenario 1.1 Values in males | 0.290 - 6.38 [1.33] | 18 | L-T NOAEL, S-T NOEL, Acute LD50, Chronic NOAEL | Miar, Apsy, Mumu, Mygl | Max, Q3, Med, Min | 22 (23) | 22 | (1) | 36 (38) | 47 | 70 | 75 | (7) |
Scenario 1.2 Values in females | 0.080 - 1.76 [0.368] | 10 | L-T NOAEL, S-T NOEL, Acute LD50, Chronic NOAEL | Miar, Apsy, Mumu, Mygl | Max, Q3, Med | 9 (10) | 9 (10) | 15 (16) | 19 (21) | 26 (30) | 31 | ||
*Values considered close to the trigger: less than 1 of difference, e.g. value of 5.2 for a trigger of 5
Importantly, in the study on small mammals carried out in the same area and mentioned above (Fritsch et al., 2022), 112 different residues of banned and currently used PPP (parent chemicals and metabolites) were detected in hair samples from wood mice and greater white-toothed shrews, with 32 to 65 residues detected per individual. This means that “cocktail effects” are likely to occur, which can ultimately increase the risk of GLY- or GLUF-induced toxicity assessed here owing to mechanisms of additivity and synergy (Martin et al., 2021). In their review, Martin et al. (2021) reported that additivity was the interaction that most often occurred in chemical mixtures, followed by synergistic interactions. For example, Brodeur et al. (2014) reported synergic toxic impacts in the common South American toad (Rhinella arenarum) of binary mixtures of GLY-and cypermethrin-based commercial PPP products; two compounds that have been detected in more than half of the small mammal individuals in our studies. They repoted that the degree of synergy ranged from 2 to 9 times depending on the mixture tested (Brodeur et al., 2014).
From terrestrial wildlife and biodiversity conservation perspectives, our findings may be worrisome. As mentioned earlier, GLY concentrations were found at higher levels in bats than in this study (Hooper et al., 2022). The authors emphasized that GLY reached the highest concentrations measured in their study, where 8 compounds, including neonicotinoids and several systemic or selective herbicides, were analysed. These authors suggested that extremely high environmental inputs of GLY may counteract the role of its short half-life in mitigating risks to wildlife (Hooper et al., 2022). A growing body of evidence raises concerns about the lethal and sublethal toxic effects of GLY in biota and highlights endocrine disruption and reproductive impairment in vertebrates chronically exposed to environmental concentrations (Fritsch et al., 2024). Recent studies have shown the toxic effects of GLUF at environmentally relevant concentrations and suggest that it has deleterious consequences on biota health under chronic exposure scenarios (Santovito et al., 2024). For example, genomic instability and cellular damage in vitro in cultured human lymphocytes and in vivo in Lymnaea stagnalis haemocytes following low-dose exposure to glufosinate-ammonium have been reported, as have negative impacts on reproductive health in L. stagnalis (Santovito et al., 2024). Furthermore, some wild taxa may be highly sensitive to GLY, AMPA or GLUF, more than expected from laboratory studies on models such as rats and mice, and commercial formulations can be more toxic than a.s. alone (Fritsch et al., 2024a). For instance, the mortality rate of postmetamorphosis juveniles of the species Rana sylvatica, Bufo woodhousii fowleri, and Hyla versicolor reached 68-86% when they were exposed via spraying to a GLY-based commercial herbicide at the recommended dose (Relyea, 2005). The exposure of spined toads (Bufo spinosus) to AMPA at environmental concentrations affects hatchling and tadpole morphology and decreases embryonic and tadpole survival (Cheron & Brischoux, 2020, 2023).
Risks of indirect effects of GLY on wildlife via disturbances in natural habitats and dietary resources due to effects on nontarget plants and arthropods [from offsite spray drift] have been noted in risk assessments (EFSA, 2015c; US EPA, 2020). Following GLY treatments, altered habitat and/or food resources have been associated with reduced (breeding) populations and negative effects on reproduction in birds (Lazaran et al., 2013; Ojelade et al., 2022), reduction in the frequentation of elk populations (Milner et al., 2013), and a reduced abundance of small mammals (especially insectivores Soricidae and herbivores Microtinae) (Santillo et al., 1989).Our results show a pervasive exposure, and they question the magnitude of positive compensatory effects (i.e. refuges and connectivity recolonization) expected from untreated areas within the agricultural landscape mosaic (see for instance Dalkvist et al. (2013) about the role of landscape features in the impacts of pesticides on vole population dynamics). Moreover, recurring treatments over seasons and years and use over large surfaces may lead to pseudopersistence within the environment (Hvězdová et al., 2018) and thus likely limit the possibility for recovery. Indirect effects may thus occur in addition to direct toxic effects.
Our findings warrant further research to confirm their interpretation and gather knowledge about the ecological impacts of the use of GLY and GLUF as PPP. There is a need to characterize the toxicokinetics and toxicodynamics of these compounds in wild species better and to provide data to relate concentrations in hair with exposure dose, accumulation in other tissues, and toxicological responses. Furthermore, more research is needed to assess whether the potential risks computed here actually translate into deleterious effects in free-ranging populations, which would require field surveys and monitoring of wildlife exposure and health. The dynamics of GLY, AMPA and GLUF transfer and their impacts on food webs crucially need to be investigated.
Here, we provide lines of evidence to support the identification of the organophosphate chemicals GLY and GLUF as ‘emerging organic contaminants’, as emphasized by Kissane and Shephard (2017). Given the pervasive exposure of wildlife to GLY, AMPA and GLUF and suggesting chronic exposure at sublethal levels on the basis of nondestructive sampling in biosentinel species, our study highlights the need for research to fill gaps in knowledge about early warning signals of impacts as highlighted by Kissane and Shephard (2017).
Conclusion
Our results provide new insights into the ecological risks of GLY- and GLUF-based herbicides, showing that in addition to the evidence related to environmental and human health issues, these PPP are of concern for wildlife exposure and may be of concern for impacts and further trophic cascading effects. The exposure of wild small mammals to GLY, AMPA and GLUF was shown to be widespread in all trophic groups or taxa and in treated and nontreated habitats within the studied agricultural area and was unrelated to the intensity of the treatments at the plot or landscape scale. These results support the most recent findings concerning pervasive GLY environmental contamination, occurrence in food webs, and associated ecological risks. Our results warrant further research on the ecological safety of the use of GLY and GLUF as PPP and provide support for assessing their potential as ‘emerging organic contaminants’.
Our study relied on measurements of residues in hair samples, a method that is developing in human epidemiology, and looks a promising nondestructive approach to study the field ecotoxicology of PPP and monitor wildlife exposure or detect early signals in wild species. Although this approach should be strengthened by further assessment of exposure doses and measurements of effects in animals, such surveys may represent valuable opportunities for the assessment of regulatory measures and postregistration monitoring and provide support for phytopharmacovigilance and toxicovigilance schemes (Vijver et al., 2017). For example, the presence of pesticides in the hair of small mammals reported in Fritsch et al. (2022) was reported as an exposure warning for nontarget species by the French department of the agency ANSES in charge of phytopharmacovigilance.
This study highlights the need for research on the pathways of wildlife exposure to PPP and the toxicokinetics and toxicodynamics of GLY, AMPA and GLUF and, more broadly, of PPP initially assessed as not persistent and not bioaccumulative on the basis of criteria established on legacy compounds. Our findings strongly suggest that current agri-environmental schemes do not achieve the objectives of ecological risk mitigation. They call for additional research and monitoring of the role of agroecological infrastructure in shaping PPP impacts on biodiversity and provide support for an overall reflection, if not an overhaul, of agroenvironmental policies.
Appendices
Supplementary Information Annex 1
Text A1: Molecular identification of small mammal species
Text A2: Reagents and procedures used for the chemical analyses of glyphosate, AMPA and glufosinate
Text A3: Approaches of reverse dosimetry to assess the risk for deleterious effects in small mammals
Text A4: Statistical outputs of the Tarone-Ware tests
Figure A1. Correlations between the concentrations of glyphosate, AMPA, and glufosinate in small mammal hair.
Figure A2. Simplified phylogenetic tree of the rodents and shrews included in the study.
Figure A3. Concentrations of glyphosate, AMPA, and glufosinate in the hair of small mammals with nondetects set to 0 according to the proportion of organic farming in the landscape as a continuous factor (a) and to the proxy of treatment intensity at township scale as a continuous factor (b) and as a categorical factor (c).
Figure A4. Probabilities of detection of glufosinate in hair of small mammals according to the sales of the active substance at the township scale.
Table A.1. Overview of sample size.
Table A.2. Statistical outputs of binomial GLMs used to investigate the influence of explanatory factors on the detection frequencies of glyphosate, AMPA, and glufosinate in the hair of small mammals.
Table A.3. Statistical outputs of the GLMs used to investigate the influence of explanatory factors on the concentrations of glyphosate, AMPA, and glufosinate in the hair of small mammals.
Supplementary Information Annex 2
Computation of reverse dosimetry approaches (calculation spreadsheets).
Acknowledgements
We thank the French National Institute INRAE and the French RECOTOX network for their financial support. We thank all of the volunteers and participants of the project “PING” for their efforts in preparing or contributing to the field and laboratory work. We especially acknowledge the help of Renaud Scheifler, Louisiane Burkart and Grégoire Perez with the field work. We thank the farmers who agreed that the sampling could be performed in their fields. We thank the PEA2t platform of the Chrono-Environnement Department, which manages the equipment used for the experiments and for molecular analyses.
Preprint version 3 of this article has been peer-reviewed and recommended by Peer Community In Ecotoxicology and Environmental Chemistry (https://doi.org/10.24072/pci.ecotoxenvchem.100211; Labadie, 2024).
Funding
This work was financially supported by the Metaprogramme SMaCH of the French INRAE (research call 2017) and the French network “RECOTOX”.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data, scripts, code, and supplementary information availability
Data are available online: https://doi.org/10.25666/DATAUBFC-2024-08-29 (Fritsch, 2024).
Scripts and code are available online: 〈hal-04485797v3〉; https://hal.science/hal-04485797 (Fritsch et al., 2024b).
Annexes are available online: 〈hal-04485797v3〉; https://hal.science/hal-04485797 (Fritsch et al., 2024b).