
 CC-BY 4.0
CC-BY 4.0
Introduction
In the majority of temperate areas, herbivore livestock are frequently raised outdoors for a portion of the year (Van Laer et al., 2014). Such outdoor rearing generally goes with grassland or rangeland use whether production objectives are meat or milk (EFSA, 2014). From the animal’s perspective, pasturing offers several advantages in terms of welfare and health (Van Laer et al., 2014; Mellor, 2015). Space allowance can reduce social aggressions and allows animals to move. Grazing allows herbivores to express exploratory and selective behaviours in diversified pastures. These benefits are consistent with the positive consumers’ perception of grass-based feeding systems (Font i Furnols et al., 2011, for the example of lamb meat purchasing intentions). However, outdoor rearing also goes with some drawbacks and risks (Temple & Manteca, 2020), from which the variability in feed quality and availability, the load of parasitism or the threat of predators are particularly important. Another great constraint is the climatic one, including in temperate areas. Under the context of climate change, with the prevision of an increase in the occurrence of extreme events (Nardone et al., 2010), the climate concern is becoming an increasing challenge for animal production (Polsky & von Keyserlingk, 2017). The challenge is particularly important for grazing herbivore livestock systems which are affected both directly via effects on animals (e.g. thermal stress) and indirectly via impacts on the resources they feed on (forage availability and quality, water availability).
Among climatic constraints, heat stress poses a significant risk and has received extensive research attention, likely attributed to its detrimental impacts on animal production (Rashamol et al., 2019). Some reviews have shed light on the negative consequences of heat stress on ruminants (biological functioning, health, welfare, production) as well as on the coping strategies at the animal and system levels to try to overcome them (Marai et al., 2007; Polsky & von Keyserlingk, 2017; Herbut et al., 2018). For animals reared outdoors, solar radiation is considered as the greatest environmental risk factor of heat stress (Herbut et al., 2018), making the provision of shade an effective solution for improving animal welfare. Positive effects of shade on animal comfort have been evidenced by changes in daily activity patterns (increased feeding time, increased rumination time in the lying posture), increased time spent in shade or lowered panting scores and body temperature, when compared to unshaded conditions as well as in relation to change in weather conditions (Tucker et al., 2008; De et al., 2020; Marcone et al., 2021). In grazing systems, trees can serve as natural sources of shade and shelter for animals. Compared to artificial shelters, trees can additionally offer several ecosystem services such as increased biodiversity, improved soil fertility and control of erosion (Torralba et al., 2016; Castle et al., 2022), making the interest for silvopastures growing, consistently with the consideration of agroecological concepts. In sheep breeding systems, there is however little research on the effects of the availability of trees and tree shade on sheep behaviour, welfare (heat stress), health and production (De et al., 2020). This is even more scarce if we look at temperate regions (Marcone et al., 2021; Pent et al., 2021), and at the active use of tree shade by sheep in the light of welfare purposes (Pent et al., 2020a).
The study aimed to objectify the importance of trees for sheep welfare by characterising how productive ewes, grazing in temperate mid-mountain pastures, use shade depending on climatic conditions and tree density. We assumed that trees would have a positive effect on ewes by alleviating heat stress and we predicted that ewes would actively and increasingly seek out the shade from trees as climatic conditions became more stressful. With the decrease in tree density, and therefore in shade availiability, we also predicted that ewes’ motivation to get shade (selectivity for shade) will become more pronounced. Additionally, as trees can negatively affect sward biomass (Hawke, 1991; Kallenbach et al., 2006), we complemented the measurements on animals with measurements on the feeding resource (pasture vegetation) to provide elements of interpretation regarding trends in animal performance.
Materials and Methods
The general procedure consisted of analysing behaviour, especially shade use, activity, welfare and performances of sheep within a silvopastoral system characterised by three tree densities. The experiment focused on the grazing season (spring and summer, May to September in our temperate area) and, for behavioural observations, on sunny days with increased thermal load on the animals. We repeated the design over three consecutive years, using different groups of sheep, in order to capture some of the variability due to variations in weather conditions (Sollenberger, 2015).
Experimental site and animals
The experimental procedures were validated by the regional ethical committee and approved by the French Research Ministry under n°2016050900291012.
The experiment was conducted in 2016, 2017 and 2018 at the INRAE Experimental Unit (UE1414 “Herbipôle”) in Central France (45°42’53”N, 3°01’21”E, 850m). The climate is temperate and semi-continental with mountainous influences and classified as Cfb in the Köppen-Geiger classification (Beck et al., 2018). The experimental setting was composed of three pastures of similar size (0.82 ha, SD 0.034) located within a radius of 320 m (Figure 1). They were pastures of permanent grasslands, composed mainly of Lolium perenne L., Holcus lanatus L., Poa sp. and Agrostis sp. as grasses, and Trifolium repens L., Achillea millefolium L., Stella sp. and Veronica sp. as legumes and forbs. The pastures were traditionally grazed by the sheep flock of the farm during the grazing season for several years before the experiment, and were not fertilized.
The three experimental pastures, named Tlow, Tmed, and Thigh were characterised by the presence of mature deciduous trees, planted in the last 1980s. These pastures comprised an increasing density of trees, from one tree in Tlow, 60 trees/ha in Tmed and 150 trees /ha in Thigh. Trees species were wild cherry (Prunus avium L.) in Thigh and Tmed, completed in Tmed by some maple trees (Acer pseudoplatanus L.), while the tree in Tlow was an ash tree (Fraxinus excelsior L.). Trees were planted within the pastures, in more or less regular rows (Figure 1). All were sufficiently high so that none of their branches nor leaves were reachable by the animals. In order to assess the percentage of tree coverage within the pastures, we used the diameter of tree crowns, measured by using satellite images at summer time then analysed with the ImageJ software and completed with in situ measurements. The calculated coverages were 0.8%, 40% and 81% for Tlow, Tmed and Thigh, respectively. They were sufficient for all ewes and their twin lambs to be in the shade at the same time, including in Tlow. Each year, a new batch of 30 Romane ewes plus their 60 lambs were used, except in 2018, when five out of the 30 had already been used in 2017. They were aged between 2 and 7 years and weighed on average at the start of the
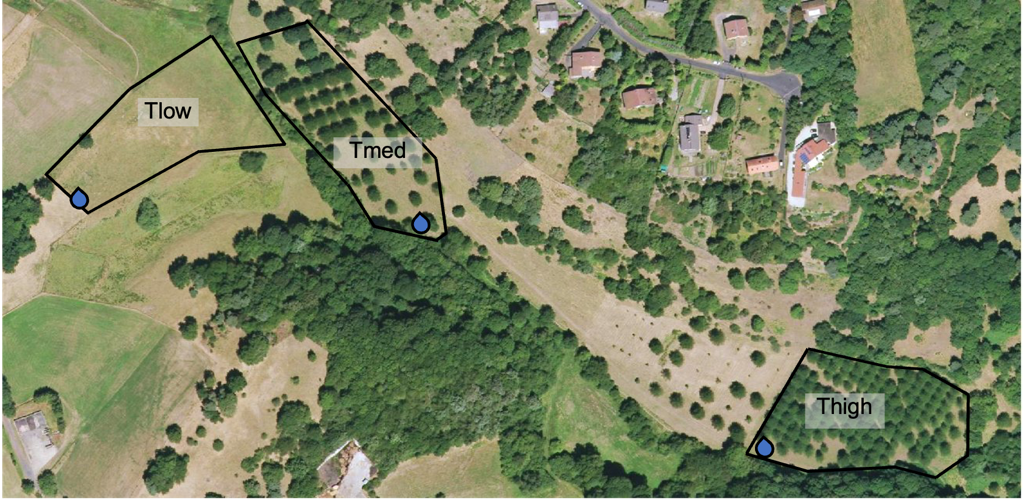
Figure 1 - Aerial view of the three experimental pastures. Tlow is the control pasture with one tree, Tmed has a tree density of 60 trees/ha, Thigh has 150 trees/ha. The blue drops indicate the position of the water supply (from Google Earth).
experiment 72.6 kg (SD 6.7) in 2016, 66.5 kg (SD 7.5) in 2017 and 67.5 kg (SD 7.8) in 2018. For the first 1.5 month of the experiment each year (from mid-May to end June), the ewes were suckling twin lambs, until those were weaned at about 2.5 months of age. To facilitate identification, the 10 animals in each group were marked with coloured stripes on their sides and back using sprays commonly used in farming. The marking was carried out for the first time in mid-May, just before they entered the experimental pastures, and was refreshed as needed during the weighing of the animals.
The ewes have been shorn once, during the first half of March, one month before lambing. In December of the previous year, three weeks after their return to the sheepbarn, the ewes were dewormed against gastro-intestinal and pulmonary strongyles (Albendazole: 15 mg/kg BW; Moxidectine: 200 μg/kg BW).
Grazing and ewe management
Each year, the thirty ewes were grouped by ten according to live weight, body condition score and age, and allocated to one of the three pastures Tlow, Tmed and Thigh. The experiment was considered to start when the ewes and their lambs were introduced into the experimental pastures, which occurred on 16th May 2016, 17th May 2017 and 17th May 2018. Before these dates, the ewes were housed indoors where they lambed around mid-April. By the end of June, the lambs were weaned and ewes temporarily returned indoors during one week for drying. Before weaning, the lambs were all day long with their mothers.
We applied a continuous grazing on the pastures and applied the rule consisting of excluding the group of ewes of a given pasture when the mean sward height on that pasture fell below 5.5 cm. Sward height was assessed every 10 to 15 days, on the basis of 200 measurements per pasture, along 10 parallel transects, using the Hill Farming Research Organisation (HFRO) sward stick (Barthram, 1986). In 2016, the grazing period ended on October 11th, September 12th and August 08th for Tlow, Tmed and Thigh groups, respectively. In 2017, the grazing period ended on September 16th, 13th and 06th for Tlow, Tmed and Thigh groups, respectively. In 2018, Tlow and Tmed groups left their pastures on August 16th, and Thigh on August 9th. At pasture, neither the ewes nor the lambs were supplemented. They all had free access to water (in water bowser or a trough, Figure 1) and salt blocks (located near the water supply) at all times.
Measurements
Animal measurements
We observed the activity of all ewes on 4 days in 2016, 8 days in 2017 and 7 days in 2018, spread between May and August each year (Table 1). We have deliberately focused on sunny days with temperatures forecast at or above the monthly average, in order to consider conditions with some risk of heat stress.
No other disturbances or measurements (animals’ weighing, sward measurements) were planned or observed on these days. Observations were made on three time slots (morning, mid-day and late afternoon) of 2 hours in 2016 and 2.5 hours in 2017 and 2018. The time windows were 8:00-11:00, 12:30-15:15, 16:30-20:00 and were adjusted according to the different day lengths in spring and summer. Observations were made simultaneously on the three groups of ewes by three different observers using the scan sampling method on a 5-min basis. The observers changed pastures evenly over the observation days (between time slots). At each scan, the observers noted the posture of each ewe (standing, lying), its activity (grazing, resting, ruminating, moving and other activities), and its position relative to shade (under shade or in the sun when sunny weather; or “no sun” when cloudy or overcast weather). A scan was considered “sunny” as soon as a shadow could be identified on the ground. An animal was considered under shade when its head or more than half of its body was under shade. Unless otherwise stated, data are presented relative to the whole day observed (all timeslots combined). During the same observation days, between scans, we recorded the respiratory rates of the ewes while they were immobile (resting or ruminating), by counting the number of flank movements over 1 to 1.5 minute.
Over the grazing period (when the ewes were on the experimental pastures), we recorded ewes’ body weight and body condition score every 2 to 3 weeks. Lambs were weighed at the same occasions as long as they were with their mothers. Animals weighing occurred on the morning and within the pastures thanks to the use of a mobile scale.
Table 1 - Climatic conditions of the observation days over the 8h-20h time slot (Mean: average of hourly measurements; Max: value from the hour with the highest value).
Date | Temp. Mean (°C) | Temp. Max (°C) | Rad. Mean (Wh/m²) | Rad. Max (Wh/m²) | Humidity (%) | WS (m/s) | TRH | THI | Prop. sunny scans |
|---|---|---|---|---|---|---|---|---|---|
2016-05-27 | 19.5 | 21.2 | 580 | 969 | 63.9 | 3.9 | 0.71 | 65.2 | 0.83* |
2016-06-06 | 18.2 | 20.6 | 577 | 941 | 73.4 | 4.1 | 0.16 | 63.5 | 0.81* |
2016-07-20 | 27.9 | 31.2 | 384 | 739 | 35.1 | 4.3 | 2.14 | 73.2 | 0.62 |
2016-07-25 | 21.9 | 23.2 | 484 | 806 | 58.3 | 4.3 | 0.86 | 68.3 | 0.78* |
2017-05-22 | 19.7 | 21.4 | 473 | 980 | 48.1 | 3.8 | 0.99 | 64.7 | 0.63 |
2017-05-23 | 18.9 | 21.2 | 491 | 935 | 72.9 | 5.2 | -0.05 | 64.7 | 0.61 |
2017-06-01 | 17.5 | 19.4 | 345 | 689 | 80.1 | 3.7 | -1.03 | 62.8 | 0.44 |
2017-06-08 | 21.5 | 24.8 | 678 | 958 | 53.6 | 3.8 | 1.71 | 67.3 | 1.0* |
2017-07-18 | 26.3 | 29.6 | 662 | 924 | 45.8 | 4.8 | 2.52 | 72.6 | 1.0* |
2017-07-28 | 18.8 | 21.4 | 583 | 958 | 65.3 | 5.5 | 0.58 | 64.1 | 0.76* |
2017-08-02 | 26.4 | 29.9 | 582 | 890 | 57.7 | 4.1 | 1.76 | 74.2 | 0.94* |
2017-08-17 | 22.9 | 25 | 584 | 874 | 56.2 | 4.3 | 1.42 | 69.3 | 0.93* |
2018-06-01 | 15.8 | 18.1 | 320 | 526 | 87.4 | 3.5 | -1.61 | 60.1 | 0.21 |
2018-06-04 | 18 | 20.3 | 501 | 806 | 75.7 | 4.3 | -0.23 | 63.5 | 0.57 |
2018-06-08 | 18.1 | 19.6 | 387 | 521 | 83.4 | 3.7 | -0.94 | 63.9 | 0.27 |
2018-07-17 | 19.2 | 21.9 | 644 | 997 | 64.8 | 6.4 | 0.87 | 64.6 | 0.83* |
2018-07-18 | 21.8 | 24.1 | 659 | 1030 | 55 | 3.8 | 1.62 | 67.9 | 0.98* |
2018-07-25 | 24.9 | 26.5 | 549 | 795 | 48.3 | 4.3 | 1.85 | 71.4 | 0.77* |
2018-07-26 | 26.7 | 28.3 | 645 | 902 | 44 | 3.3 | 2.58 | 73 | 1.0* |
Temp. = Ambient temperature; Rad. = Radiation; WS = Wind speed; TRH = synthetic climatic parameter from PCA involving temperature, radiation and humidity; Prop. sunny scans = proportion of total daily scans at which animals could be noted under shade or in the sun;* = days selected for analyses of shade use and selection; THI1 formula from Mader et al., 2006.
THI=(0.8*ambient temp)+(((% relative humidity)/100)*(ambient temp-14.4))+46.4
Pasture measurements
We harvested sward samples for assessment of biomass and pasture quality at two occasions during the grazing period each year: by the end of May (spring season) and the end of July (summer season). At each date, we harvested 16 samples per pasture, along W-shaped transects. For Tmed and Thigh pastures, eight samples were harvested in open areas and eight samples in areas under tree crowns. For Tlow, all the samples were harvested in the open area (n=8 in 2016 and n=16 in 2017 and 2018). Each sample corresponded to a 0.1x2m strip of sward using a hand mower cutting at 2 cm from soil surface. Samples were then dried at 60°C for 72h in a ventilated oven before being grinded for subsequent chemical analysis. Sward samples were analysed using near infrared spectroscopy (NIRS) to determine contents of crude protein (CP; AOAC, 1990), of neutral detergent fibre (NDF; Van Soest et al., 1991), and pepsin-cellulase dry matter digestibility (dCell; Aufrère & Michalet-Doreau, 1983). Near infrared spectra were collected with a monochromator (FOSSNIRSystems 6500, Silver Spring, MD, USA), which scans between 400 and 2500nm every 2 nm. For CP, the global calibration model obtained by Andueza et al. (2011) was used. For NDF and dCell, models obtained by Andueza et al. (2016) were used.
Climate monitoring
To characterise the local climatic conditions, we used the monitoring data from the INRAE CLIMATIK platform (https://agroclim.inrae.fr/climatik) managed by the AgroClim laboratory of Avignon, France (Delannoy et al. 2022). The selected weather station (n°63345002; 45°43’23”N, 3°01’10”E) is located at about 750m from the central point of the three pastures, at 890m of altitude. From this station, we accessed hourly data of ambient temperature, radiation, humidity and maximal wind speed. From these data, we selected the 8h-20h time slot to characterise the observation days (Table 1).
In order to analyse the behaviour of ewes in relation to climatic conditions, we aimed to synthesise the different climatic parameters (temperature, radiation, humidity and wind speed) into one value. Various thermic stress indices exist in the literature but none has been developed in agroforestry conditions (see discussion). Similarly to Stachowicz et al. (2019), we thus considered the four main climatic parameters cited above, over the 8h-20h time slot for all the days of the grazing period each year, and ran a principal component analysis (PCA) on these variables to derive a synthetic parameter for the climatic characterisation of days (see Statistical Analysis section below).
The INRAE CLIMATIK weather station also allowed us to characterise the grazing months (May to September) of the three experimental years in terms of temperature, radiation, humidity, rainfall and wind speed (Table 2).
Table 2 - Climatic characteristics of the three experimental years, from May to September (mean (standard deviation)). Mean: monthly average of daily mean measurements (8-20h); Max/Min: monthly average of daily maximal/minimal measurements.
Year | Month | Temp. Mean (°C) | Temp. Max (°C) | Temp. Min (°C) | Rad. Mean (Wh/m²) | Rad. Max (Wh/m²) | Hum. Mean (%) | WS Mean (m/s) |
|---|---|---|---|---|---|---|---|---|
2016 | May | 12.4 (3.7) | 14.5 (4.0) | 9.6 (3.6) | 409 (169) | 696 (239) | 67.9 (12.7) | 7.1 (2.4) |
June | 15.9 (4.0) | 17.9 (4.2) | 13.1 (3.4) | 409 (153) | 717 (214) | 73.7 (9.7) | 5.4 (1.8) | |
July | 19.9 (4.4) | 22.3 (4.5) | 16.6 (4.0) | 494 (154) | 815 (165) | 63.3 (16.3) | 5.0 (1.2) | |
August | 21.2 (4.9) | 23.9 (5.0) | 17.3 (4.6) | 468 (137) | 786 (159) | 53.1 (16.2) | 5.0 (1.0) | |
Sept. | 18.0 (4.6) | 20.5 (5.1) | 13.6 (3.8) | 336 (119) | 617 (170) | 64.3 (18.7) | 4.7 (1.6) | |
2017 | May | 14.7 (5.7) | 16.8 (5.8) | 11.7 (5.6) | 448 (171) | 760 (206) | 71.2 (16.2) | 5.7 (1.8) |
June | 19.4 (4.9) | 21.7 (5.0) | 16.3 (4.7) | 492 (161) | 809 (164) | 71.6 (13.3) | 5.3 (2.4) | |
July | 19.6 (4.1) | 21.9 (4.6) | 16.1 (3.3) | 470 (145) | 768 (185) | 69.2 (12.5) | 5.4 (1.6) | |
August | 20.5 (5.0) | 23.0 (5.5) | 16.8 (4.1) | 446 (138) | 729 (183) | 63.9 (14.0) | 4.7 (1.3) | |
Sept. | 13.5 (3.4) | 15.6 (3.7) | 10.1 (2.5) | 326 (103) | 634 (178) | 74.9 (10.5) | 5.4 (2.6) | |
2018 | May | 13.7 (4.1) | 16.1 (4.3) | 10.9 (4.4) | 374 (149) | 714 (187) | 78.3 (10.9) | 5.3 (1.6) |
June | 17.7 (2.9) | 19.7 (3.0) | 14.7 (3.0) | 500 (169) | 790 (187) | 74.4 (11.0) | 4.7 (1.1) | |
July | 21.3 (3.0) | 23.7 (3.0) | 18.0 (3.0) | 532 (132) | 860 (149) | 61.3 (10.5) | 4.7 (0.9) | |
August | 20.9 (4.4) | 23.2 (4.5) | 17.3 (3.9) | 486 (127) | 808 (138) | 59.7 (11.9) | 5.3 (1.5) | |
Sept. | 18.2 (4.2) | 21.1 (4.6) | 13.2 (3.7) | 395 (122) | 683 (170) | 59.5 (14.2) | 4.5 (1.7) |
Temp.= Ambient temperature; Rad.= Radiation; Hum.= Humidity; WS= Wind speed.
Statistical analyses
In order to calculate a synthetic parameter for the climate characterisation of the observation days, we ran a PCA with the R software (R Studio 4.2.3) using the FactoMineR package (Lê et al., 2008). We used a total of 364 days over the three years and considered the climatic parameters over the 8h-20h time slot. The final PCA involved temperature, radiation and humidity, which were well represented on the 1st dimension and explained 77% of the variance (Kaiser-Meyer-Olkin score, KMO=0.70). As wind speed was the only variable represented on the second dimension, it was considered on its own, as an explanatory variable, for subsequent analyses. For the rest of the document, the 1st dimension of the PCA (i.e. the coordinates of each day on this dimension) will be named TRH value (for temperature, radiation, humidity). For interpretation, as TRH value increases, climatic conditions move towards higher temperature and radiation and lower humidity. These TRH values for observations days are presented in Table 1. The TRH values of all days as well as the outputs of the PCA are presented in Appendix 1 and 2.
We performed the other analyses with the SAS® Enterprise Guide (7.1 version) software, completed with the XLSTAT (2020.3.1) software for non-parametric analyses when conditions for parametric ones were not satisfied.
Data on animal behaviour were the proportion of time spent under shade (shade use), the selection of shade for the activities of grazing and resting + ruminating (i.e. the proportion of a given activity that was observed under shade = shade selection), and the proportion of time spent
standing while resting or ruminating. For these data, only days with at least 75% sunny scans were considered. The 12 selected “sunny” days (out of the 19) were evenly distributed over the 3 years (Table 1). Regarding shade use and shade selection data, only the sunny scans within these days were considered. Regarding the proportion of time spent standing, all scans within these days were considered. These data were analysed in relation to the climatic characterisation of observation days (TRH and wind speed), using the Mixed Procedure of SAS®. Data were occasionally submitted to transformation to improve the distribution of residuals. This occurred for data relative to shade selection for resting + ruminating activities, and to proportion of standing time, which were subjected to the arcsine transformation. The model tested the fixed effects of treatment (Tlow, Tmed, Thigh), TRH, wind speed and their interactions. The square value of TRH (so TRH²) was also included to test for the apparent curvilinear shape of the relation between animal behaviour and TRH. We included as random effect the group of ewes nested within year (as different groups were used each year), with the variance components covariance structure. The statistical unit was the group of ten ewes (average values of the ten animals). The difference between treatments was assessed both globally and for specified values of TRH (0, 0.5, 1, 1.5, 2 and 2.5) by pairwise comparison corrected by the Tukey adjustment method. Regarding shade use and shade selection specifically, we performed post-hoc one-sample Wilcoxon signed-rank tests to compare, for each treatment, the proportion of time ewes spent under shade with the proportion of pasture area covered by tree canopy. This aimed to assess whether ewes from each treatment were actively searching tree shade while shade provision differed according to tree density. In addition, shade use was characterised by calculating the Jacobs’ Selectivity Index (Si, Jacobs, 1974), which allows assessing the selectivity for a ressource taking into account its availability in the environment. The formula is:
Si=(ci-ai)/(ci+ai-2ciai )
where ai is the proportion of pasture area covered by tree canopy, used as a proxy of shade availability, and ci is the proportion of time spent under shade by the ewes. The index ranges from -1 (total avoidance) to +1 (total selection), with 0 indicating that the resource is used in proportion of its availability. We used the non-parametric Kruskall-Wallis test to compare treatments and assess the ewes’ motivation to use shade as its availaibility decreased from Thigh to Tlow. We carried out the post-hoc one-sample Wilcoxon signed-rank and the Kruskall-Wallis tests on daily data and at the specific midday time slot when the sun's position minimises the shadow cast by the surrounding area.
Data on animal performances (ewes’ body weight and body condition score, lambs’ body weight) were analysed by year, on individual animals, using the Mixed Procedure of SAS® with the Repeated statement to account for the individuals being measured at different dates, associated with the autoregressive covariance structure. Regarding lambs, the analysed data is the average of twin lambs weight. The effects tested were the treatment, the weighing date and their interaction. The random effect was the individuals nested within treatments. To account for multiple pairwise comparisons, the p-values were considered after correction by the Tukey adjustment method. Regarding ewes’ performances, data included the weighing dates as long as at least two out of the three treatments were still on the experimental pastures (see “Grazing and ewe management” section).
Data on respiratory rates were firstly averaged per animal and per observation day, then per year so as to obtain one value per animal and per year. As the conditions for parametric analysis were not met, we used the Kruskall-Wallis test to analyse the effect of treatments by considering all years together.
Data on vegetation (biomass, CP and NDF contents, dCell) were analysed per year, using the Mixed Procedure of SAS®. We tested the effect of treatment, season and their interaction, with the consideration of the repeated statement to account for that sampling occurred in spring and summer. CP data were subjected to log transformation. As for the other analyses, the pairwise comparisons were corrected by the Tukey adjustment method.
For the mixed models analyses, effect size estimates and their standard errors are presented in Appendix 3. Additionally, eta square (η²) values for the Kruskall-Wallis tests are provided within the main text.
Results
Animal behaviour
Time under shade (shade use) and shade selectivity
The proportion of time the ewes spent under shade was affected by the treatment (p < 0.0001) and climatic ambiance (TRH, p = 0.0085), but not by their interaction (p = 0.20) nor by wind speed (p = 0.10) (Figure 2). Square TRH was significant (p = 0.046). Neither the interaction of treatment with either wind speed or TRH² was significant, and these interactions were removed from the model. Overall, the analysis indicates that the proportion of time spent under shade decreased from Thigh to Tmed then Tlow (Tmed—fixed effect estimate ± SE = -0.196 ± 0.07, p = 0.013; Tlow-fixed effect estimate ± SE = -0.504 ± 0.07, p < 0.0001; Table S1 Appendix 3).
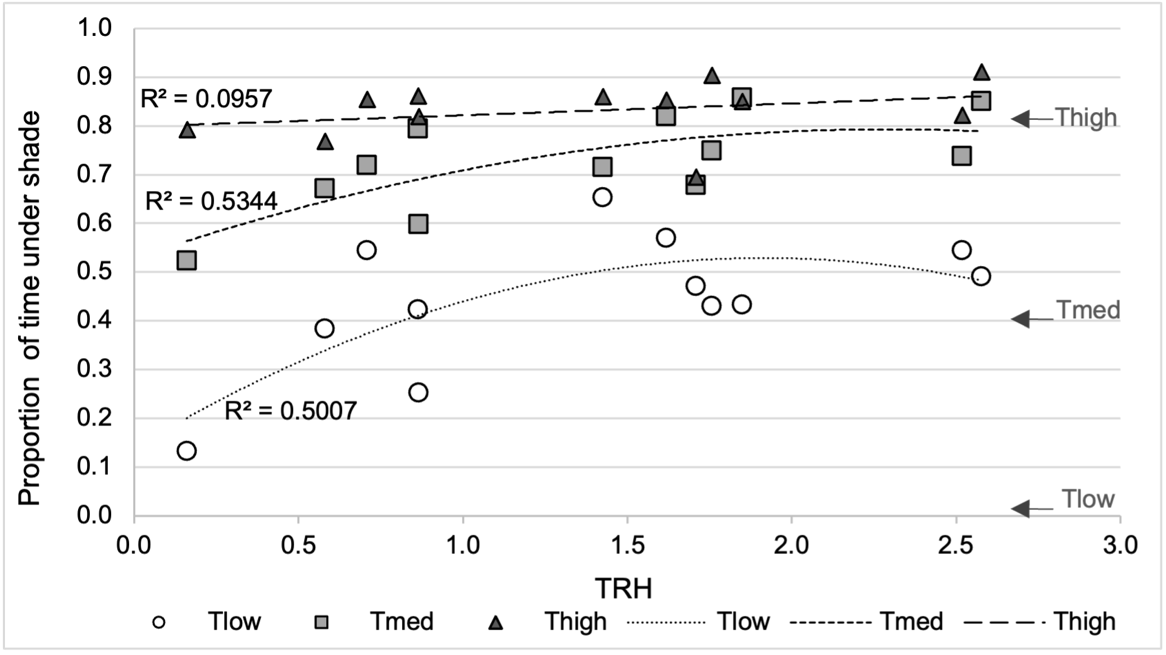
Figure 2 - Proportion of time spent under shade by the ewes depending on tree density (Tlow: one tree, Tmed: 60 trees/ha, Thigh: 150 trees/ha), and the synthetic climatic parameter TRH (Temperature, Radiation, Humidity). Each point represents the average proportion for a group of 10 ewes, over the three experimental years (12 sunny days). The lines represent the curvilinear trends for each treatment (with the R² coefficients associated). The arrows on the right represent the surface of the pastures covered by tree crowns (0.8% in Tlow, 40% in Tmed, 81% in Thigh).
Despite the non-significance of the treatment*TRH interaction, the analysis of least squares mean differences at specified TRH values indicates a similar shade use for Tmed and Thigh treatments at TRH values of 2.0 and 2.5, whereas all three treatments differed from each other at the lower specified TRH values. For all treatments, the proportion of time spent under shade increased with TRH (TRH-fixed effect estimate ± SE = 0.169 ± 0.08, p = 0.048; Table S1 Appendix 3). The significance of TRH² suggests the trend of a curvilinear fit for the evolution of time spent under shade relative to TRH (TRH²-fixed effect estimate ± SE = -0.055 ± 0.026, p = 0.046; Table S1 Appendix 3).
The one-sample Wilcoxon signed-rank tests indicate that for both the daily and midday time slots, the mean shade use was greater than the tree canopy cover for Tlow and Tmed, in contrast to Thigh where it was not different (Table 3).
Table 3 - Mean and median of shade use (proportion of sunny scans spent under shade) over all sunny days (n=12) according to time slots and treatments, and p-value of one-sample Wilcoxon signed-rank tests comparing shade use with the theoretical value of tree canopy cover (proportion of pasture surface).
Treatment | Mean | Std Dev | Median | Tree canopy cover | p-value | |
|---|---|---|---|---|---|---|
Day (3 time slots) | Tlow | 0.44 | 0.14 | 0.45 | 0.008 | 0.0005 |
Tmed | 0.73 | 0.10 | 0.73 | 0.40 | 0.0005 | |
Thigh | 0.83 | 0.06 | 0.85 | 0.81 | 0.18 | |
Midday time slot | Tlow | 0.43 | 0.22 | 0.49 | 0.008 | 0.001 |
Tmed | 0.71 | 0.15 | 0.72 | 0.40 | 0.0005 | |
Thigh | 0.74 | 0.10 | 0.74 | 0.81 | 0.055 |
Tlow: one tree, Tmed: 60 trees/ha, Thigh: 150 trees/ha
The Jacobs’ index of selectivity (Si ) differed between treatments for both the daily (p < 0.0001, η² = 0.76) and midday time slots (p = 0.0001, η² = 0.48 ) (Figure 3). The Tlow ewes expressed the greatest Si and the Thigh ewes the lowest values. All treatments differed from each other at the day scale. At midday, the Tlow and Tmed treatments did not appear to differ, due to two specific days with a use of shade by some or all of the Tlow ewes very different (much lower) from all the other days of observation.
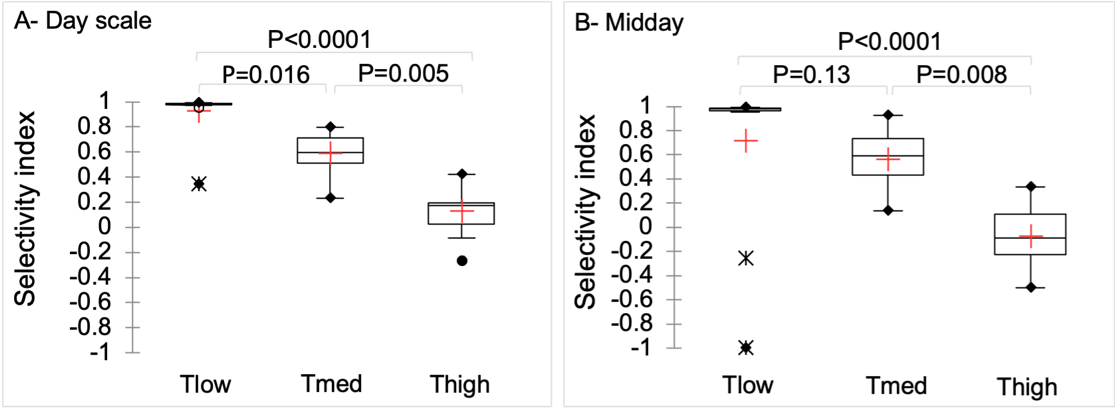
Figure 3 - Jacobs’ selectivity index for shade use at the day scale (A) and for the midday time-slot (B), depending on tree density (Tlow: one tree, Tmed: 60 trees/ha, Thigh: 150 trees/ha). Within box plots, the red cross represents the mean, and the traits the median and the 1st and 3rd quartiles. Whiskers extend to 1.5 times the interquartile range and data out of this range are plotted individually.
Shade selection for main activities
As a preliminary step to looking at shade selection for the main activities, we analysed the proportions of time spent on these activities, i.e. feeding, resting and ruminating. Over the twelve experimental days (all scans), these proportions were similar between treatments (p = 0.92, p = 0.21 and p = 0.34, respectively). Ewes spent an average of 58% (SD 9.6) of the observed scans feeding, 20% (SD 6.8) resting and 19% (SD 7.7) ruminating.
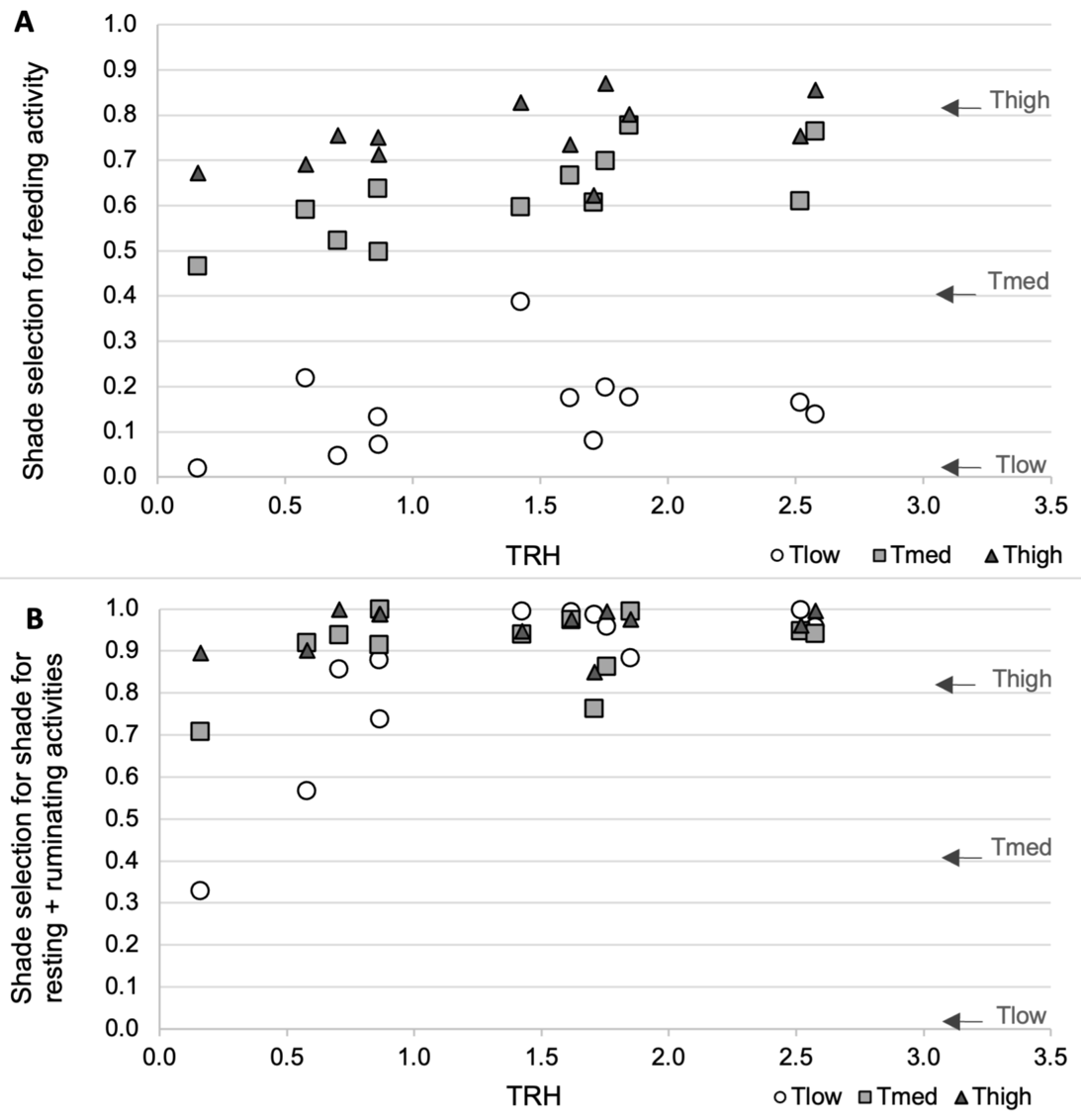
Figure 4 - Shade selection (proportion of time of a given activity spent under shade) by the ewes for feeding (A), and resting + ruminating (B) activities depending on tree density (Tlow: one tree, Tmed: 60 trees/ha, Thigh: 150 trees/ha), and the synthetic climatic parameter TRH (Temperature, Radiation, Humidity). Each point is the average proportion for a group of 10 ewes, over the three experimental years (12 sunny days). The arrows on the right represent the surface of the pastures covered by tree crowns (0.8% in Tlow, 40% in Tmed, 81% in Thigh).
Regarding the feeding activity, the selection of shade decreased from Thigh to Tmed then Tlow (treatment effect, p < 0.0001; Tmed—fixed effect estimate ± SE = -0.196 ± 0.07, p = 0.014; Tlow-fixed effect estimate ± SE = -0.588 ± 0.07, p < 0.0001; Table S2 Appendix 3) and also increased with TRH (p = 0.017; TRH-fixed effect estimate ± SE = 0.159 ± 0.08, p = 0.053; Table S2 Appendix 3) for all treatments. No other effect was significant (Figure 4A). Relative to tree canopy cover, the Tlow and Tmed ewes actively selected shade for the feeding activity (one-sample Wilcoxon signed-rank tests, p = 0.0005) whereas the Thigh ewes avoided it (p = 0.03).
Regarding the sum of resting and ruminating activities, the selection of shade was affected by the treatment (p = 0.007), TRH (p = 0.0006), their interaction (p = 0.006) and TRH² (p = 0.005) (Figure 4B). The fixed effect estimates, on the basis of the arcsine transformation of this variable, are as follows: Tmed—fixed effect estimate ± SE = -2627 ± 8669, p = 0.77; Tlow-fixed effect estimate ± SE = -29500 ± 8669, p = 0.004; TRH-fixed effect estimate ± SE = 26185 ± 8365, p = 0.004 (Table S2 Appendix 3). At the TRH specified values of 0 and 0.5, shade selection was lower in Tlow than Tmed and Thigh ewes, then lower in Tlow than Thigh ewes at TRH value of 1 (p < 0.05). These differences were no longer visible at greater TRH specified values for all comparisons (p > 0.05). The ewes from Tmed and Thigh never differed in their selection of shade. We can also notice that from TRH values of 1 and above, almost all shade selection data were between 0.8 and 1 with several ones at 1 (all treatments represented) indicating a frequent exclusive selection of shade for resting and ruminating activities. Accordingly, the one-sample Wilcoxon signed-rank tests indicate that the ewes from all three treatments were actively selecting shade for resting and ruminating (p = 0.0005 for Tlow, Tmed and Thigh).
Proportion of resting and ruminating time spent standing
The proportion of resting and ruminating time spent standing was not affected by any of the effects tested (treatment, TRH, treatment*TRH, wind speed, TRH², p > 0.2) (Figure 5). We observed only a tendency for a treatment effect once all other effects were removed from the model (p = 0.08), but the fixed effect estimates show no significant effect (Table S3 Appendix 3).
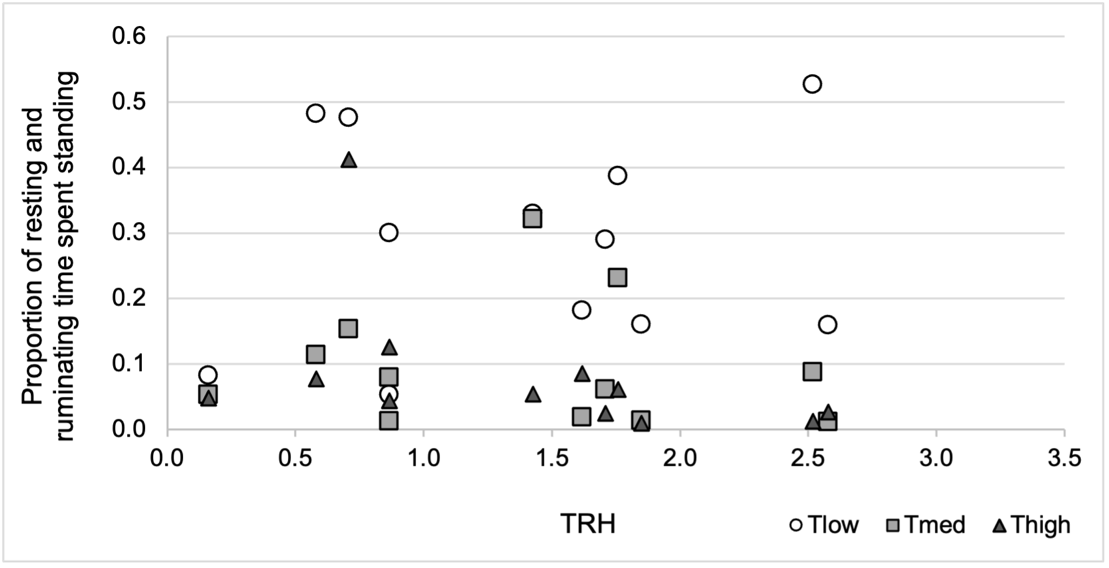
Figure 5 - Proportion of resting and ruminating time spent standing, depending on tree density (Tlow: one tree, Tmed: 60 trees/ha, Thigh: 150 trees/ha), and the synthetic climatic parameter TRH (Temperature, Radiation, Humidity). Each point represents the average proportion for a group of 10 ewes, over the three experimental years (12 sunny days).
Respiratory rate
Over the three experimental years, the respiratory rate was affected by the treatment (Kruskall-Wallis test: K = 15.76, p = 0.0004; Figure 6), but the effect size is low (η² = 0.16). The greatest rate was recorded in Tlow ewes, while Tmed and Thigh ones showed similar rates.
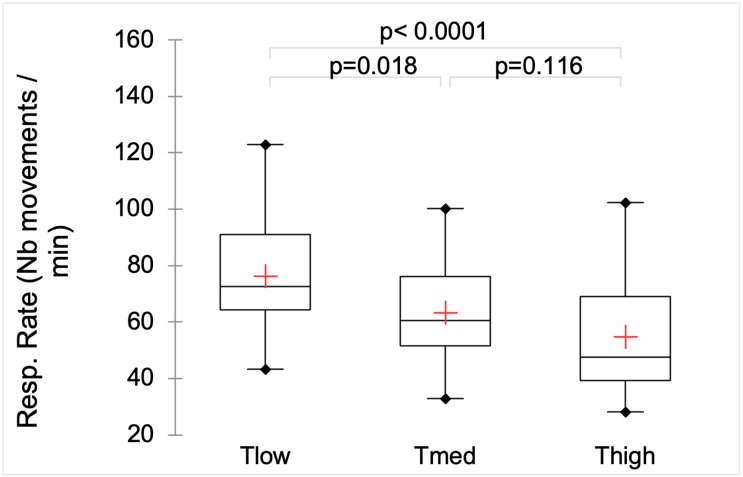
Figure 6 - Effect of tree density (Tlow: one tree; Tmed: 60 trees/ha; Thigh: 150 trees/ha) on ewes’ respiratory rate (number of flank movements per minute) for the all three experimental years. Within box plots, the cross represents the mean, and the traits the median and the 1st and 3rd quartiles. Whiskers extend to 1.5 times the interquartile range and data out of this range are plotted individually.
Table 4 - p-values of the tested effects relative to performance data for ewes and lambs
Data | Effect | 2016 | 2017 | 2018 |
|---|---|---|---|---|
Ewes’ Body weight | Date | 0.0001 | 0.0001 | 0.0001 |
Treatment | 0.26 | 0.95 | 0.27 | |
Date*Treatment | 0.0001 | 0.0016 | 0.0001 | |
Ewes’ Body condition score | Date | 0.0001 | 0.0001 | 0.0001 |
Treatment | 0.17 | 0.46 | 0.34 | |
Date*Treatment | 0.027 | 0.0001 | 0.015 | |
Lambs’ body weight (average weight of twin lamb) | Date | 0.0001 | 0.0001 | 0.0001 |
Treatment | 0.95 | 0.42 | 0.40 | |
Date*Treatment | 0.11 | 0.0005 | 0.029 |
Animal performance
Ewes’ body weight (BW) and body condition score (BCS)
At all years and for all treatments, the ewes lost BW over the grazing season, from -10% in 2018 for Tlow ewes to -25% in 2016 for Thigh ewes, between the first and the last weighing (Table 4, Figure 7, Table S4 for the fixed effect estimates). The general trend in 2016 and 2017 was a decrease in BW until the post-drying weighing, followed by either a maintenance or a re-increase afterwards. The pattern of 2018 was slightly different with a more progressive decrease in BW all along the grazing period on pastures without re-gaining at the end. The shorter duration of the grazing period in 2018 may have participated in this different trend. The differences between treatments are expressed in interaction with the weighing dates, with an advantage for the Tlow treatment compared to the other two treatments. In 2016, Tlow ewes gained body weight after drying (D59-D122, p = 0.0001) while Tmed ones did not; in 2018, Tlow ewes maintained their body weight from D13 up to the end while Tmed and Thigh ewes lost body weight during the same period (Tmed: p = 0.0075, Thigh: p = 0.0001). Nevertheless, these differences never led to significant differences between treatments at the different weighing dates, after p-values were corrected for multiple comparisons. The trends for BCS were comparable to those of BW (Table 4, Figure 7, Table S5 for the fixed effect estimates), excepted that the Tlow ewes in 2016 and the ewes from all treatments in 2017 had recovered their pre-grazing BCS by the end of the period on experimental pastures. In 2016, the Tmed and Thigh ewes had lost BCS at the end of the period (Tmed: -0.8 point, p = 0.0003; Thigh: -0.9 point, p = 0.0001, compared to Tlow ewes: -0.3 point, p = 0.9), as well as the ewes from all treatments in 2018 (Tlow: -0.7 point, p = 0.0002; Tmed: -0.9 point, p = 0.0001; Thigh: -0.5 point, p = 0.05). As for BW, BCS never differed between treatments at the different scoring dates.
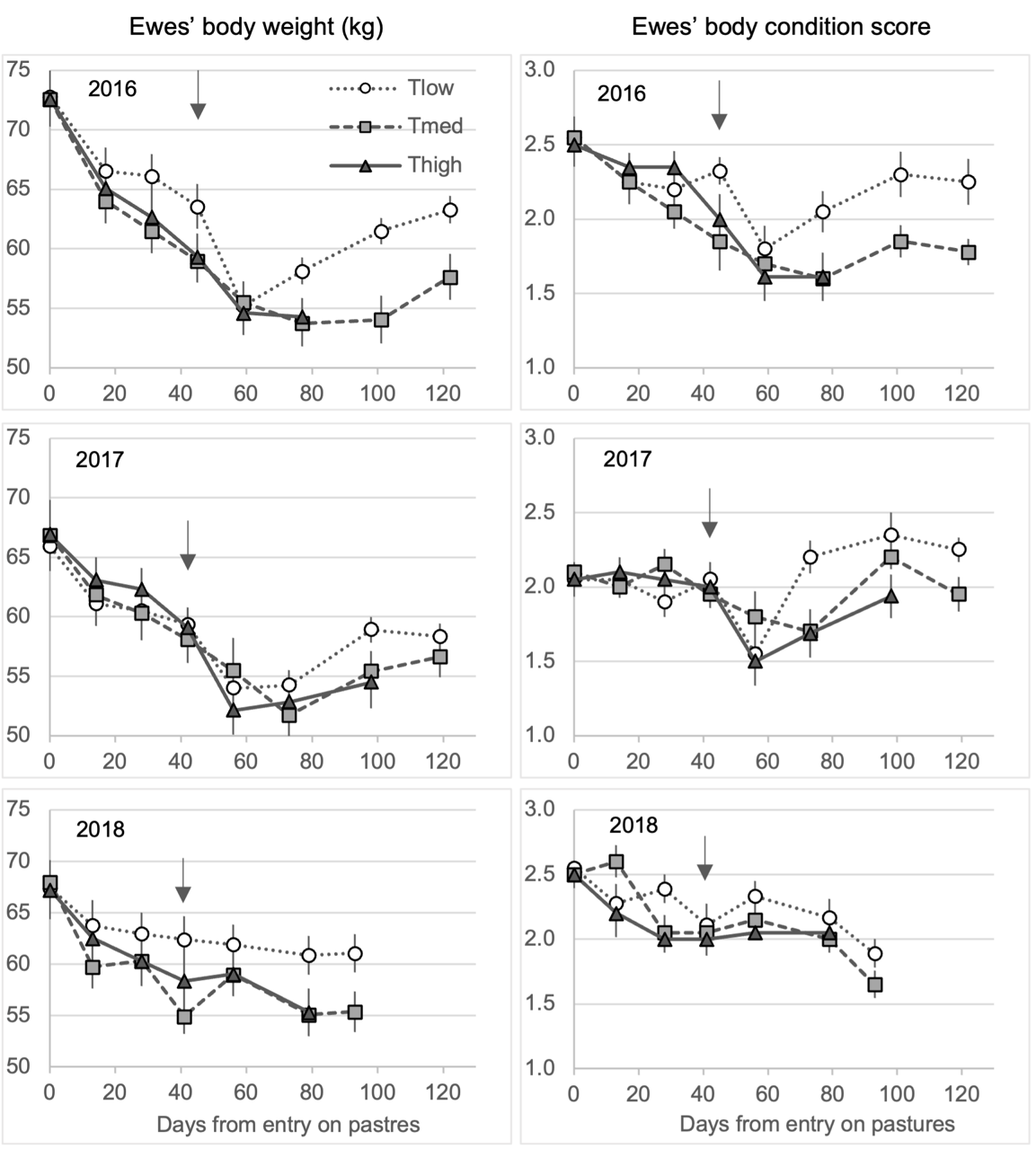
Figure 7 - Evolution of ewes’ body weight (kg) and body condition score (scale of 0 to 5) (mean ± standard error) from all treatments, according to the number of days since the entry on pastures, each year. The number of days differed between years and treatments (see text for explanation). Data are presented as long as at least two treatments were on the pastures. The white circles represent the Tlow treatment (one tree), grey squares the Tmed treatment (60 trees/ha) and black triangles the Thigh treatment (150 trees/ha). The arrows represent the time of lambs weaning.
Lambs’ body weight (BW)
Whatever the year and treatment, all lambs’ weights (average of twin lambs’ weight) increased quite linearly over the weighing dates (Table 4, Figure 8, Table S6 for the fixed effect estimates). In 2017 and 2018, the date*treatment interaction was significant but once the correction for pairwise comparisons applied, no difference was found between treatments regardless of the date.
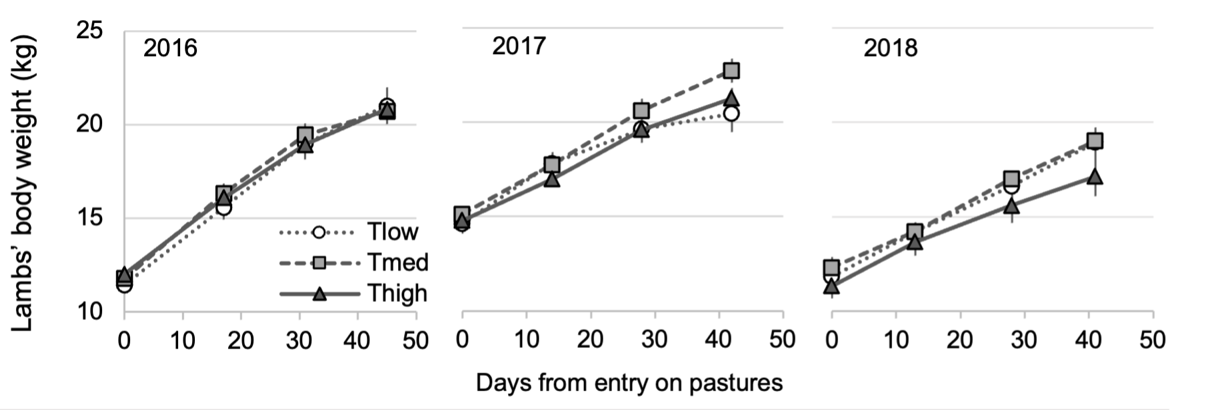
Figure 8 - Evolution of average of twin lambs’ body weight (kg) (mean ± standard error) from the three treatments, according to the number of days since the entry on experimental pastures, for the three experimental years. The white circles represent the Tlow treatment (one tree), grey squares the Tmed treatment (60 trees/ha) and black triangles the Thigh treatment (150 trees/ha).
Pasture characteristics
Biomass
For all three years, the sward biomass was higher in Tlow than in Tmed and higher in Tmed than in Thigh, with a decrease of 50% on average between Tlow and Thigh (treatment effect, p = 0.0001 each year) (Figure 9, Table S7 for the fixed effect estimates). As the season progressed, the biomass increased in 2016 (from spring to summer, p = 0.0001), but decreased in 2017 (p = 0.004) and 2018 (p = 0.0001). The treatment*season interaction was never significant.
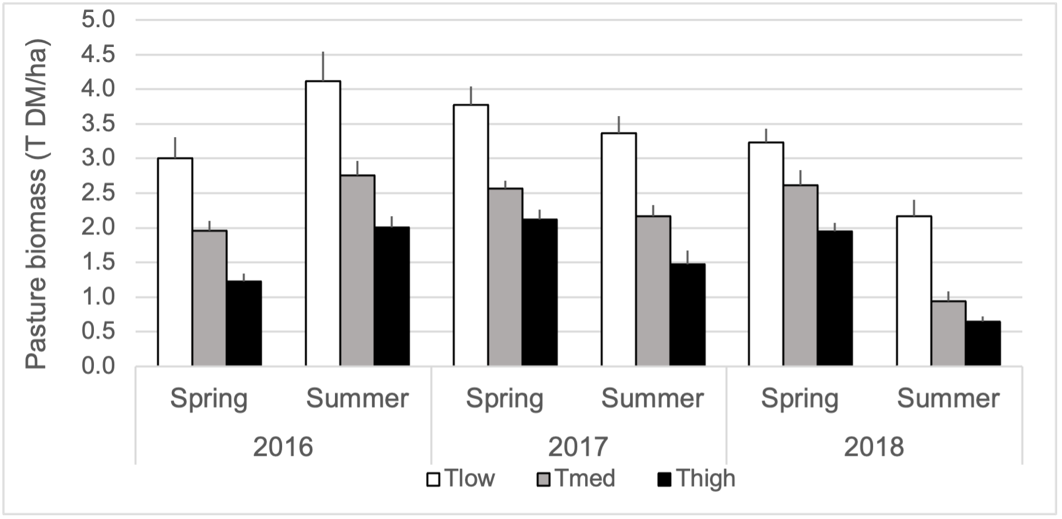
Figure 9 – Pasture biomass (mean ± std error, tonnes of dry matter per hectare) according to tree density (Tlow: one tree; Tmed: 60 trees/ha; Thigh: 150 trees/ha) and season, over the three experimental years and seasons.
Chemical composition
The CP content of the pastures was greater in Thigh and Tmed compared to Tlow in 2016 and 2017 (Treatment effect: p = 0.0004 and p = 0.0001, respectively) (Table 5, Table S7). In 2018, the same pattern was observed in summer while no difference was observed in spring (treatment*season effect: p = 0.0015). This interaction also revealed a decrease in CP content between spring and summer 2018, but only in Tlow. The decrease in CP content with season was also observed in 2016 (p = 0.0001), while no difference was evident in 2017 (p = 0.7).
Table 5 - Pasture quality (CP: crude protein; NDF: Neutral detergent fibre; dCell: Cellulase digestibility; mean and standard error) according to treatment (Tlow: one tree; Tmed: 60 trees/ha; Thigh: 150 trees/ha), year and season.
CP (g/kg DM) | NDF (g/kg DM) | dCell (Proportion DM) | ||||||||||
Tlow | Tmed | Thigh | Tlow | Tmed | Thigh | Tlow | Tmed | Thigh | ||||
Mean | ||||||||||||
2016 | Spring | 106 | 115 | 115 | 575 | 571 | 564 | 0.623 | 0.612 | 0.539 | ||
Summer | 82 | 109 | 102 | 597 | 578 | 544 | 0.458 | 0.474 | 0.410 | |||
2017 | Spring | 93 | 108 | 104 | 563 | 557 | 588 | 0.603 | 0.619 | 0.584 | ||
Summer | 85 | 115 | 113 | 644 | 640 | 634 | 0.424 | 0.451 | 0.444 | |||
2018 | Spring | 107 | 124 | 120 | 606 | 588 | 609 | 0.523 | 0.548 | 0.533 | ||
Summer | 93 | 148 | 128 | 634 | 568 | 632 | 0.441 | 0.525 | 0.423 | |||
Standard error | ||||||||||||
2016 | Spring | 4.5 | 3.9 | 3.9 | 8.3 | 9.1 | 10.8 | 0.018 | 0.010 | 0.019 | ||
Summer | 4.6 | 5.4 | 3.2 | 14.5 | 7.0 | 10.8 | 0.018 | 0.013 | 0.014 | |||
2017 | Spring | 2.2 | 5.9 | 2.7 | 7.2 | 7.7 | 8.1 | 0.014 | 0.013 | 0.016 | ||
Summer | 1.5 | 6.1 | 5.3 | 4.6 | 9.9 | 7.2 | 0.008 | 0.022 | 0.018 | |||
2018 | Spring | 2.2 | 3.9 | 4.4 | 6.3 | 7.8 | 5.8 | 0.015 | 0.013 | 0.015 | ||
Summer | 3.4 | 9.1 | 6.8 | 8.9 | 8.1 | 10.8 | 0.013 | 0.021 | 0.018 | |||
The NDF content was lower in Thigh than in Tlow in 2016 (Treatment effect, p = 0.02). In 2017 and 2018, we observed a treatment*season interaction (p = 0.03 and p = 0.007, respectively). Once considered the correction for multiple comparisons, no difference between treatments was observed in 2017 nor in spring 2018. In summer 2018, the lowest NDF content was observed in Tmed compared to Tlow and Thigh, which did not differ from each other. The NDF content remained stable between seasons in 2016 (p = 0.74) and 2018 (p > 0.15 for all treatments). In 2017, the NDF content increased between spring and summer (p < 0.0005 for all treatments).
Regarding dCell, the differences between treatments changed depending on the year (Table 5). Digestibility was lower in Thigh than in Tmed and Tlow in 2016 (treatment effect, p = 0.0001), similar between all treatments in 2017 (p = 0.3) as well as in spring 2018. In summer 2018, dCell was greater in Tmed than in the other two treatments (2018: treatment*season effect, p = 0.03). The evolution through seasons was more consistent, with a decrease in digestibility between spring and summer for all treatments in 2016 and 2017 (p = 0.0001 for both years), and for Tlow and Thigh treatments in 2018, while no evolution was observed for the Tmed treatment.
Discussion
The consideration of the climatic constraints imposed on livestock reared outdoors is a growing concern. For those animals, the main environmental risk factor of thermal stress is solar radiation (Herbut et al., 2018). Hence, the provision of shade can be viewed as a critical point for the welfare of livestock, especially during hot and sunny days. In order to argue for the provision of shade, it is crucial to objectively assess not only the effects of providing shade on animal welfare or performance, but also the use of shade by animals in different climatic conditions. Our experiment aimed to provide data for this objectification in the specific context of sheep grazing temperate permanent pastures with varying tree densities, during sunny days in spring and summer. Our main hypothesis was that ewes would actively seek out tree shade, due to its mitigating effect on heat stress, and that this would intensify as climatic conditions became more stressful. With the decrease in tree density, and therefore in shade availiability, we also predicted that ewes’ motivation to get shade (shade selectivity) will become more pronounced.
The study, based on very contrasted tree densities and a follow-up of three years, provides a comprehensive overview and valuable insights into the effects of trees on sheep in grazed pastures. Its limitations lie in the limited number of observation days, which focused only on sunny weather, the rough assessment of shade provision between treatments, which was approximated by the pasture area covered by tree crowns, and the observations made by several observers. Within these limitations, the main results align with the formulated hypotheses, and lay the groundwork for more detailed future research.
Characterisation of observation days in terms of risk of heat stress
Heat stress occurs when the environmental conditions, particularly temperature, rise and reach a level that triggers the activation of defence mechanisms to maintain homeothermy (Silanikove, 2000). The presence, magnitude or risk of occurrence of heat stress are generally assessed either by some indices based on climatic data or by data measured directly on the animals (Herbut et al., 2018).
Various indices exist in the literature that integrate two or more climatic parameters to provide one assessment of the climatic ambiance declined as thermic stress or heat load, with increasing risk levels according to increasing thresholds: e.g. Temperature humidity index (THI, Thom, 1959; Mader et al., 2006), adjusted Temperature humidity index (THIadj, Mader et al., 2006) , Heat load index (HLI, Gaughan et al., 2008), Black globe humidity index (BGHI, Buffington et al., 1981) or Comprehensive climate index (CCI, Mader et al., 2010). These indices and their subsequent evolutions are well presented in some reviews such as the one of Herbut et al. (2018) or Wang et al. (2018). According to the equations developed in the papers cited above, our observation days (12 sunny days, 8h-20h time-slot, climatic parameters assessed from the weather station positioned in open area without shade) ranged between 63.5-74.2 for THI, 61.2-75.5 for THIadj_hourly, 65.8-81.3 for HLI, 71.2-81.8 for BGHI and 14.6-26.1 for CCI. When we consider the thresholds presented for these various indices in the literature, the number of our observation days that would fall within the thermal neutral or comfort zone of the animals, is 11 for THI, 10 for THIadj, 4 for HLI and 10 for CCI. For BGHI, 4 out of the 12 days are in the neutral zone if THI thresholds are applied (Wang et al., 2018), but none would be considered “normal” in the conditions stated by Herbut et al. (2018).
Wang and collaborators (2018) emphasized the importance of carefully considering the conditions under which the index is developed (type of environment and animal, assumptions, modelling method) so as to prevent any misuse or misinterpretation. The target species for the development of most of the heat stress indices are not sheep but cattle (Rashamol et al., 2019). In sheep, most studies used THI and to a lesser extent BGHI. In the specific sheep studies that examined shade-related concerns, THI was selected (e.g. Sevi et al., 2001; Pent et al., 2020a), although THI does not consider solar radiation nor wind speed, because of its development for cattle in a confined environment type (Wang et al., 2018). All these elements led us to characterise and classify our days for the analysis of ewes’ behaviour relative to the climatic conditions, independently from the existing indices. As in Stachowicz et al. (2019), we used a synthetic climatic parameter (TRH), derived from the local climatic data recorded during the grazing periods over the course of the three experimental years. TRH values correlate with higher temperature and radiation but lower humidity, consistent with the characteristics of the local temperate climate with mountainous influences. This contrasts with tropical climates where higher humidity is a risk factor for greater heat stress.
Nevertheless, for the purpose of comparing and discussing our results with previous studies using THI, it is useful to position our days of observation relative to this index (Table 1). On the basis of the thresholds identified for cattle (Mader et al., 2006), our conditions appeared not stressful (THI < 74), or only mildly so.
An active use of shade by the ewes
The primary goal of this study was to investigate how ewes, given varying degrees of shade availability, would use the shade in relation to climatic conditions. Information is scarce about sheep responses, including their behaviour, in environments providing shade and about the implications on their welfare (De et al., 2020). Only a few recent studies have explored how sheep use shade (Ginane et al., 2018; Maia et al., 2020; Pent et al., 2020a; Leu et al., 2021; Marcone et al., 2021), typically offering only shaded and non-shaded treatments without varying shade availability. A slightly larger number of studies have compared the main activities and/or physiological responses of sheep in shaded versus non-shaded environments (e.g. Sevi et al., 2001; De et al., 2020; Pent et al., 2021). This highlights the need for data on sheep behaviour in relation to shade. Our main finding is that the proportion of time spent in shade by ewes increased both with tree density and to a lower extent with the synthetic climatic parameter (TRH).
The increased use of shade with increasing shade provision is consistent with previous studies on cattle, which involved varying degrees of shade (number of m²/animal or % of pasture shaded), through the use of artificial or natural shelters (Schütz et al., 2010; Rosselle et al., 2013; Schütz et al., 2014). The greater shade use with tree density is logical as the probability for the animals to be in shade mathematically increases with shade provision. The interesting point is to know whether they actively seek shade and to what extent they are motivated to obtain it.
The active search of shade was analysed by comparing, for each treatment, the proportion of time spent under shade with the proportion of shade available on the pastures. We estimated shade availability by the areas covered by tree crowns, resulting in percentages of 0.8, 40 and 81% for Tlow, Tmed and Thigh pastures, respectively. In the Thigh pasture, the proportion of time ewes spent in shade was similar to the shade availability (83% on average). In contrast, for pastures with lower tree density, the proportion of time spent in the shade was significantly higher than its availability: 73% versus 40% in Tmed, and 44% versus 0.8% for Tlow. These marked differences clearly show an active search for shade, even though the shade provision assessment did not account for the changing shadow casts due to the sun’s orientation throughout the day. Focusing on the midday time slot, which minimises the underestimation of shade availability, confirms this active search of shade in Tmed (71% of time spent in shade) and Tlow (43%). These high levels of shade use support the reports of some previous works on shade preference, motivation for and active seeking of shade in cattle (Bennett et al., 1985; Schütz et al., 2008) and sheep (Pent et al., 2020a), in order to alleviate heat stress.
To delve deeper into motivation, behavioural studies typically assess motivation for a resource by restricting its availability, as done from Thigh to Tlow. If shade is important, we predict an increase in shade selectivity (selection relative to availability) as its availability decreases. We measured this using the Jacobs’ index of selectivity (Jacobs, 1974), traditionnaly used for analysing animal diet selectivity (e.g. Dumont et al., 2007), but also applied in other contexts (e.g. Barros & Pereira, 2014). The results support the predictions, showing an increase in selectivity from almost no selection in Thigh (Si=0.12) to very high selection (Si=0.95) in Tlow, on a daily scale. This is confirmed at midday (Si=-0.08 for Thigh, Si=0.71 for Tlow) although we observed more variability between days at this time scale than on the daily scale in Tlow ewes.
A selection of shade primarily for resting and ruminating activities
The ewes from all three treatments also increased their proportion of time spent in shade with the synthetic climatic parameter (TRH). Despite the lack of interaction between treatment and TRH, the increase logically appears stronger in the treatments with medium and low tree density. For both treatments, and particularly for Tlow, we observe a plateau, while the ewes were actively seeking shade. It is therefore interesting and informative to examine for what activities the ewes used shade. The selection of shade for the main activities indicates that shade was used primarily for resting and/or ruminating. This is consistent with the observations made on sheep grazing in orchards in North of France (Ginane et al., 2018). From TRH values close to 1.5, representing a theoretical day with an average temperature of 23°C, radiation of 590 Wh/m² and humidity of 56%, it can be seen that the ewes from all treatments selected shade for resting and ruminating in the same way and at very high levels, most values of shade selection being between 90 and 100%. On the other hand, the feeding activity was constrained by the availability of feed under the tree(s), especially in Tlow, due to trampling and the presence of faeces under the only available tree. This explains the lower levels of shade selection for this activity compared to resting and ruminating. Nevertheless, the selection for shade for grazing activity increased with TRH in all treatments, suggesting that consideration should be given to providing shade in a way that allows animals to graze in shaded areas when climatic conditions become restrictive.
An active shade use even on lower TRH days that may indicate thermal discomfort and risk of heat stress
The increase in proportion of time spent in shade by the ewes with increasing TRH, is consistent with previous studies on cattle, which have found an increase in shade use with increasing solar radiation (Tucker et al., 2008), ambient temperature (Rosselle et al., 2013) or microclimatic indices (THI, HLI; Kendall et al., 2006; Veissier et al., 2018). In sheep, regardless of whether the shade was provided naturally (Leu et al., 2021; Marcone et al., 2021) or artificially (Maia et al., 2020), and regardless of the study location being in a tropical (Maia et al., 2020), arid (Leu et al., 2021), or temperate area (Marcone et al., 2021), the overall results align with those obtained in studies on cattle. In the temperate area, with similar climatic conditions to ours, the use of shade by ewes, measured by the number of individuals observed under shade, showed a positive correlation with air temperature and a negative one with air humidity (Marcone et al., 2021). Therefore, there is a good consistency between these findings and our own results, as the increase in our synthetic climatic parameter (TRH) reflects rising temperatures accompanied by slight increasing radiation and decreasing humidity.
The climatic conditions of temperate regions have been less considered compared to tropical, arid or Mediterranean ones, likely partly due to the perception that they are less at risk of thermal stress for livestock. As exposed above, our observation days were classified as no or low stressful in terms of heat load (THI basis). Consistently, some of the behavioural or physiological responses that are frequently observed as an expression of heat stress, such as a lower rumination time or a greater standing time (Polsky & von Keyserlingk, 2017; Marcone et al., 2021), were no or only slightly expressed by the Tlow ewes. Despite this, we observed significant use of shade, particulalry for resting and ruminating activities. This suggests that under our temperate conditions, which are considered low-stressful, ewes may experience some thermal discomfort, if not stress, which shade helped alleviate. An increase use of shade by ewes on certain days, as we observed with rising TRH, may indicate a risk of heat stress if shade is not longer accessible. Monitoring physiological signs of heat stress (such as increased respiratory rate and panting score) on those days is important to ensure the ewes’ condition does not deteriorate.
In our study, the recorded respiratory rates were slightly higher in Tlow ewes compared to Tmed and Thigh ones, with average values of 76, 63 and 55 breaths/min, respectively. These values, along with their variations, fall within the range of no to mild stress (Silanikove, 2000; Marcone et al., 2021), but they are partly the result of the use of shade by the ewes. Without a “no shade” treatment, we can only infer the alleviating effect of shade on thermal comfort as the primary motivation for using shade. This assumption is well-supported by previous studies in sheep (De et al., 2020; Marcone et al., 2021).
A somewhat negative impact of trees on ewes’ performances
The overall pattern of evolution of ewe performances during the grazing periods showed a decrease in body weight and body condition until after weaning of the lambs, followed by either a further decrease, maintenance or recovery, depending on the treatment and year. The effect of the treatments was thus mainly manifested in this second phase (after weaning), with varying but somewhat negative effects of trees on ewes’ recovery of condition after drying. It was in 2016, and to a lesser extent in 2018, that the differences were most evident. If we compare Tlow and Tmed treatments, which involved the ewes spending the most extended period of time in the pastures, it is observed in 2016 that the Tlow ewes regained condition after drying while Tmed ones did not; in 2018, the Tlow ewes better maintained their body weight than the Tmed ones. This led to a weight loss of 13% in Tlow compared to 21% in Tmed ewes over the grazing period in 2016, and of 9% compared to 18% in 2018. However, in the absence of supplementation, all the ewes ensured good and similar growth for their lambs regardless of year and treatment.
As the ewes were not supplemented, these trends have to be considered mainly in the light of the sward characteristics. The main difference in sward characteristics between treatments was sward biomass. Biomass was impaired in Tmed and Thigh pastures compared to Tlow, whatever the year and season. In spring, the loss was about -30% in Tmed and -50% in Thigh compared to Tlow. In summer the loss was even greater, being about -40% in Tmed and -60% in Thigh but resulted from the additional effects of trees and animals’ grazing. The impairment of forage productivity due to the presence of trees is consistent with previous results. Pent et al. (2020b) and Fannon et al. (2019) related lower sward biomasses in black walnut silvopastures compared to open pastures (between -15% and -30%) at tree densities varying from 36 to 250 stems/ha depending on the study. Similarly, a decrease in pasture production up to -80% was observed in pine silvopastures as tree density increased from 0 to 200 stems/ha (Hawke, 1991). However, these reduced sward biomasses in wooded pastures did not lead to a generalised decline in animal production compared to open pastures. Results instead indicated no difference in liveweight gains (heifers: Kallenbach et al., 2006; lambs: Pent et al., 2020b) or a non-systematic decrease (Fannon et al., 2019, year effect) in silvopastures. The concurrent improvement of sward quality alongside reduced biomass participated in explaining these results (Kallenbach et al., 2006; Pent et al., 2020b). In our study, we similarly observed an overall improvement of CP content and a concomitant decrease in NDF content of Tmed or Thigh pastures compared to Tlow, but these evolutions did not seem sufficient to counterbalance the reduced sward biomasses and to allow similar ewes’ body weight and condition.
In the previously cited studies, if animals’ gains were comparable in open and in silvopastures, there were instances when the number of supported animals or animal.days over the studied period was decreased in wooded treatments (Kallenbach et al., 2006; Pent et al., 2020b). This is consistent with the systematic lower number of grazing days we observed along with the increase in tree density in our study. This was particularly noticeable in 2016, with 29 and 64 fewer grazing days in Tmed and Thigh compared to Tlow, while the difference in the other two years was limited to a maximum of 10 days. This phenomenon seems to stem from Tlow, with a remarkably high number of grazing days in 2016 (148 days) and a low number in 2018 (91 days). This may be explained by that 2016 was the only year in which biomasses increased from spring to summer to reach the maximum value recorded in the study (4 T/ha). For the same season in 2018, sward biomass barely exceeded 2 T/ha. Regarding sward quality, we did not test a year effect but no clear differences appeared between summers of 2016 and 2018 for Tlow (slightly lower CP and NDF contents in 2016) and sward quality probably did not play a significant role in these results.
Optimal tree cover
Our experiment relied on pastures and ewe flocks of limited size, with mature trees scattered within the pastures. Due to this limited flock size, one tree was sufficient to provide shelter for all ewes and their lambs at the same time. Given the high use of shade by the animals, providing sufficient shade for all individuals appears to be the primary consideration for tree cover, to avoid social competition and exclusion of some animals from access to shade (Schütz et al., 2010).
In Tlow, the ewes were constrained by the low availability of shade and limited their use of shade to resting and ruminating while they had to feed in the sun. In Tmed, ewes were able to choose to graze in the shade and took advantage of this opportunity when climatic conditions deteriorated. Thus, from a welfare point of view, several scattered trees appear to be beneficial, firstly by allowing each individual to choose a shaded or unshaded area for each activity, while remaining close to its counterparts, and secondly by spreading trample and manure across the pasture and making the sward in shaded areas grazable. On the other hand, the productivity of the pasture and the performance of the ewes were impaired in Tmed compared to Tlow. Therefore, in our conditions, an appropriate tree density would probably have been between Tlow and Tmed (e.g. 30 trees/ha) to allow ewes to benefit from tree cover while limiting the risk of negative effects on performance and pasture biomass.
Silvopastoral systems are complex and there is a lack of knowledge to understand the interactions between the different components (Jose et al., 2019), and therefore to make generalisations about optimal tree arrangements (De-Sousa et al., 2023). The design and management of a silvopastoral system regarding trees, animals and pasture impact its spatial and temporal heterogeneity (Jose et al., 2019) due to variation in pasture growth and quality and pasture use by the animals. Depending on the objectives, climate, animal species and herd size, the optimal tree arrangement may vary.
Conclusion
This study showed, in the spring and summer conditions of a temperate and mid-mountainous area, that ewes made a significant and active use of the shade provided by the trees. This active seek of shade increased with increasingly challenging climatic conditions but occurred even on days considered at low or no risk in terms of heat stress. These observations complete and confirm previous results obtained in other ruminant species, breeds and contexts, indicating a genericity in that shade is a need and contributes to the well-being of grazing livestock.
The shade-seeking behaviour was particularly highlighted at low tree density when the proportion of time spent in the shade vastly exceeded the proportion of the pasture that was shaded. With very little shade available, the ewes prioritised their activities in the shade for resting and ruminating, whereas the medium tree density provided more comfort by allowing them to choose shade for other activities as well. However, increased tree density had come with penalized sward biomass and animal performances. Further research is needed to provide data that will help determining the appropriate tree type (high canopy trees, grazable trees and shrubs, or a combination of these) and layout (isolated trees, clumps in hedgerows), as well as pasture management (continuous or rotational grazing) to best balance these positive and negative effects at the animal and system level, depending on the objectives of the system. Regarding animal behaviour, gaining a deeper comprehension of the animal’s motivations for shade, in relation with the other motivations for feed and social interactions, and their potential conflicts, in such environments will be crucial in making these determinations within the context and goals at hand.
Finally, this experiment focused on sunny days to investigate the use of shade provided by trees. To disentangle the animals’ motivation for shade from their motivation for trees for other purposes - such as shelter from rain and wind, access to certain feed compounds, scratching or hiding – additional data across various environmental conditions are required.
Acknowledgements
The authors warmly thank the staff from the INRAE Experimental and Research Units for their assistance in management and measurements on animals and swards. They also thank the trainees involved in the project: C. Gava, P. Dechavanne and M. Jardillier. B. Meunier is acknowledged for the calculation of tree canopy areas. The INRAE CLIMATIK platform from AgroClim laboratory (Avignon, France) is acknowledged for the provision of all the climate data. Preprint version 4 of this article has been peer-reviewed and recommended by Peer Community In Animal Science (https://doi.org/10.24072/pci.animsci.100303; Nielsen 2025).
Funding
This work was funded by ADEME (The French Agency for Ecological Transition) as part of the “REACCTIF 2” call for proposals.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflict of interest in relation to the content of the article.
Data, scripts and supplementary information availability
Data, scripts and summplementary information (appendices) are available online through Data INRAE (https://doi.org/10.57745/ISGTTK; Ginane et al., 2025).
Author contributions
CG: Conceptualization, Methodology, Formal analysis, Investigation, Writing – Original draft, Writing – Review & Editing. MB: Conceptualization, Methodology, Investigation, Resources. VD: Conceptualization, Methodology, Investigation, Writing – Review & Editing. DA: Resources, Writing – Review & Editing. CB: Project administration, Funding acquisition, Writing – Review & Editing.