
 CC-BY 4.0
CC-BY 4.0
Introduction
Survival and reproductive success are the two major components of Darwinian fitness, and as with all life-history traits, they are under strong selective pressures. Nevertheless, “Darwinian demons” (Law, 1979) do not exist, as species cannot evolve to live forever and reproduce continuously due to physiological and energetic constraints that create trade-offs between life-history traits (Healy et al., 2019). Yet, dietary shifts may ease energetic constraints, which could in turn allow organisms to optimise multiple fitness traits simultaneously (Swanson et al., 2016).
One striking case of dietary innovation is provided by the pollen feeding Heliconius butterflies (Gilbert, 1972; Young & Montgomery 2020). Butterflies typically require water and sugars during adulthood, which can be acquired either from rotten fruits (fruit-feeders) or nectar produced by flowers (nectar-feeders) (Krenn, 2008). Butterflies of the Heliconius genus differ from other nectar-feeders due to their ability to additionally collect and digest pollen while feeding on nectar (Gilbert, 1972; Young & Montgomery 2020). Although many insects can eat pollen (e.g. bees as well as some beetles, sawflies, mirids, thrips, flies and moths) (Wäckers et al., 2007), Heliconius are the only butterflies known to actively collect and digest pollen grains. This is probably explained by the necessity of specific adaptations for mechanical and chemical digestion of pollen to make its nutrients available for absorption (Johnson & Nicolson, 2001).
A number of adaptations were probably necessary to allow Heliconius to digest pollen. The pollen grains collected in the elongated proboscis of this butterflies are humidified with salivary secretions, aided by the co-option of a “grooming behaviour” (coiling and uncoiling of the proboscis for some minutes to hours) (Gilbert, 1972; Krenn et al., 2009; Hikl & Krenn, 2011). Moreover, a cocoonase enzyme has been found in the saliva proteome and transcriptome of Heliconius butterflies during feeding, which likely enhance pollen gemination and access to its nutrients (Harpel et al. 2015; Smith et al., 2018). Comparative transcriptomics (Smith et al., 2016) and genomics of heliconiines (Cicconardi et al., 2023) have unravelled the evolution of pollen-feeding in Heliconius and offer the possibility for the identification of other genes associated with their unique adaptation to digest pollen.
Pollen-feeding is not observed in other genera of the Heliconiini tribe and arose in the Heliconius genus, with an independent lost in the aoede clade (four species that were previously classified as the Neruda genus) (Turner, 1976; Beltrán et al., 2007; Kozak et al., 2015; Cicconardi et al., 2023). As Heliconius is the most speciose genus of the tribe, their novel ability to use pollen has likely contributed to their diversification, opening new niches to be exploited (i. e. through habitat partitioning, foodplant preference, foraging behaviour) and providing them with the energetic resources necessary for the maintenance of complex traits (Estrada & Jiggins, 2002; Montgomery et al. 2016). Indeed, the pollen-feeding behaviour of Heliconius butterflies has been associated with several aspects of their biology that diverge from the other heliconiine genera, including a long adult-lifespan (Dunlap-Pianka et al. 1977), prolonged fertility (Boggs et al., 1981; O’Brien et al., 2002), enlarged mushroom-bodies (Montgomery et al., 2016), foraging site fidelity (Moura et al., 2022) and increased adult toxicity (Pinheiro de Castro et al., 2020).
Lepidopterans generally acquire most, if not all, of their nutrients during larval feeding. By supplying butterflies with amino acids, pollen feeding may have decoupled this partition, providing a mechanism for further investment in adult behavioural strategies. Indeed, while most Lepidoptera tend to live relatively long lives as larvae and shorter lives as adults, Heliconius adults can live up to 6 months, which is much longer than the regular average life-span of other heliconiines (~1 month; Brown, 1981) despite a similar larval period (Hebberecht et al., 2022). However, when deprived of pollen, it is reported that Heliconius charithonia die at a similar average life-span to Dryas iulia, a non-pollen feeding heliconiine (Dunlap-Pianka et al., 1977). This suggest that the nutrients found in pollen contributes to Heliconius adults to invest in more durable morphology and maintaining their metabolic rates for a longer period than possible with a sugar diet alone.
Alongside this increased longevity, Heliconius butterflies also maintain their fecundity for longer than other heliconiines, as males and females show limited evidence of reproductive senescence, unless deprived of pollen (Dunlap-Pianka et al., 1977). This prolonged fertility is energetically costly: a female butterfly can lay up to 9-18 eggs a day and they can live for many months, such that total resources allocated to oviposition exceed their own body mass. Indeed, (O’Brien et al., 2003) used isotopic labelling to demonstrate the direct transfer of essential amino acids from pollen ingested by females to their eggs. Males also contribute to the cost of fertility by transferring nuptial gifts to the female during mating (Cardoso et al., 2009), which can exceed 5% of male body weight, and pollen resources may also be used for this purpose. Although the relation between diet, body weight, fertility and longevity seems obvious, there is a lack of empirical data about how pollenfeeding affects weight maintenance and how is this associated with the prolonged fertility of these butterflies.
Finally, the evolution of pollen feeding has also been associated with toxicity: Heliconius butterflies tend to have higher total concentration of cyanogenic glucosides (CG) than other heliconiines (Pinheiro de Castro et al., 2019; Sculfort et al., 2020) and mature adults have higher concentrations than larvae and young adults (Nahrstedt & Davis, 1983; Pinheiro de Castro et al., 2020). This is unusual in aposematic butterflies, which normally acquire their chemical defences from plants during larval feeding and therefore have more toxins as final instar larvae (Nishida, 2002). Whereas larvae of Heliconius balance between CG biosynthesis and sequestration from their obligatory Passifloraceae hostplants (Pineheiro de Castro et al., 2021), adults can only biosynthesize these defence compounds, for which they need the amino acids valine and isoleucine. It has been hypothesized that Heliconius butterflies use the essential amino acids from pollen for CG biosynthesis (Nahrstedt & Davis, 1983). However, studies comparing between the CG content of young Heliconius butterflies fed only sugar and supplemented with amino acids/pollen did not show any significant differences (Nahrstedt & Davis, 1985; Cardoso & Gilbert, 2013). This suggests that Heliconius butterflies might biosynthesize CGs initially using amino acids acquired during larval stage, with resources from pollen-feeding only used later in their adulthood.
Here, we explore how a dietary novelty can ease energetic constraints on life-history traits, using pollen-feeding Heliconius as a case study. We investigate the effect of pollen-feeding on H. erato body weight, chemical defences, and fertility controlling for sex and age, and specifically comparing young adults (14d) with mature adults (45d). We therefore tested the hypothesis that mature butterflies that only had access to sugar during adulthood would have lower fertility, body weight and depleted chemical defences (Boggs, 2009).
Materials and methods
Rearing conditions of H. erato stock population
All experiments were performed using individuals from a stock population of H. erato demophoon kept at University of Cambridge. This population was sourced from city of Panamá (Panama) and have been kept under insectary conditions for about 7 years. Adults were kept in breeding cages (60x60x90 cm) containing plants of Passiflora biflora for oviposition, as well as flowering Lantana sp. and few Psiguria sp. for adult feeding. Cages were provided with artificial nectar made from 10% sucrose solution (m/v) with 1.5% (m/v) Vetark Critical Care Formula (CCF). P. biflora shoots with eggs were collected from the breeding cages and used to set up larval cages (30x30x40 cm). Larvae were fed with fresh P. biflora shoots ad libitium until pupation. Larval cages were checked every other day and encountered pupae were transferred to pupal cages (30x30x40 cm), where pupae were hanged under a stick covered with a microfiber cloth. Freshly emerged individuals in the pupal cages were transferred to breeding cages. All cages were kept at 25-28°C, 60-80% humidity and 12h day/night cycle.
Experimental Design and Diet treatments
Recently emerged adults (0-1 day after eclosion) were transferred to the experimental cages (60x60x90 cm). Only adults that had morphologically healthy and dry wings were used in these experiments, with forewing between 3.0 to 3.5 cm to control for size effects. One experimental cage was set for each treatment (diet/age) and each had initially 8 males and 8 females (N=16). Butterflies that died during the first week of experiment were replaced to control for density. Butterflies were placed on feeders when added into the experimental cages to ensure that they would be able to find their food source. Each experimental cage received one of the diet treatments: 1) three feeders with artificial nectar made of 10% sucrose (m/v); or 2) three feeders with artificial nectar made of 10% sucrose (m/v) + 1.5% artificial amino acid supplement (m/v) (Vetark Critical Care Formula); or 3) three feeders with artificial nectar made of 10% sucrose (m/v) and freshly collected Lantana flowers, as a natural source of pollen. Butterflies were fed ad libitium, with feeders and flowers replaced every other day. Experimental cages were kept for 14 days to assess the importance of nitrogen on young butterflies and for 45 days to assess this effect on mature butterflies. All other heliconiines live for ~1 month, therefore 45 days is the beginning of an adulthood period that is specific to mature Heliconius butterflies. All experimental cages were kept at the same environmental conditions used for husbandry (25-28°C, 60-80% humidity and 12h day/night cycle).
Fertility assays
At the end of the experiments, female butterflies were individually assayed for oviposition to evaluate the effect of the diet treatments on fertility, while males were kept in the experimental cages until sample collection. For the fertility assays, female butterflies were transferred into individual cages (30x30x40 cm) containing their previous diet (one feeder per cage, with one flower bouquet for the pollen treatment) and a P. biflora cutting with 5 expanded leaves for oviposition. After 48h of assay, eggs were counted, weighed and collected for further analyses.
Sample collection, metabolite extraction and HPLC-MS conditions
The weight of each butterfly was recorded at the end of the experiment (14 days or 45 days). 8 males and 8 females of freshly emerged butterflies (unfed, after 0-1 day of eclosion) were also weighed and collected as a baseline. Afterwards, butterflies were collected in 1 mL methanol 80% (v/v) for chemical analyses. All samples were kept at -20 °C until further processing. For the metabolite extraction, butterfly samples were homogenized (in 1mL methanol 80% (v/v)) using a porcelain mortar and pestle. Egg samples were homogenised in 300 µL methanol 80% (v/v) into their own collection tube, using a small pestle. Extracts were centrifuged at 14,000 g for 5 min, filtered (45 µm) and collected for analyses in a LC-Orbitrap-MS/MS. LC-MS methods and analyses were conducted as described in (Pinheiro de Castro et al., 2019). The de novo biosynthesized CGs linamarin, lotaustralin and epilotraustralin were quantified in the analysed samples, which had no other CGs. The absolute amount of each compound in each sample was calculated using the peak area of their sodium adduct applied to a regression curve stablished using pure standards. Raw chemical data as well as quantification methods can be found in Pinheiro de Castro (2023).
Statistical analyses
Statistical analyses and plots were performed in R. Shapiro-Wilko test was used to analyses if the variables were normally distributed (Table S1) and Levene’s test for the homogeneity of the variances (Table S2). ANOVA was used to evaluate the effect of diet, age and sex, as well as their interaction, on butterfly weight (Table 1, Table S3 for females only). Tukey HSD was used for pairwise comparisons between the different diet:age treatments in males and females. ANOVA was used to examine the effect of diet and age on CG per laid egg with Tukey HSD for pairwise comparisons. Kruskall-Wallis was used on variables that were not normally distributed: to analyse the effect of age and diet on laid eggs; and the effect of diet, age and sex on butterfly CG content.
Results
Females are more affected than males by the lack of pollen
Age, diet and sex significantly influenced the body weight of H. erato butterflies (Table 1). There was also a significant interaction between age and sex, which indicates that the body weight of males and females were differently affected through adulthood (Table 1). Indeed, females were overall heavier than males and they were more affected by the absence of nitrogen on their adult diet (Figure 1). Mature females lost weight during adulthood without access to nitrogen, but adult diet did not affect the weight of mature males (Figure 1). Mature females that had access to pollen maintained their weight and were heavier than mature females fed sugar only, who lost weight (Figure 1). Males and females eclosed with similar weight (0d) (Figure S1).
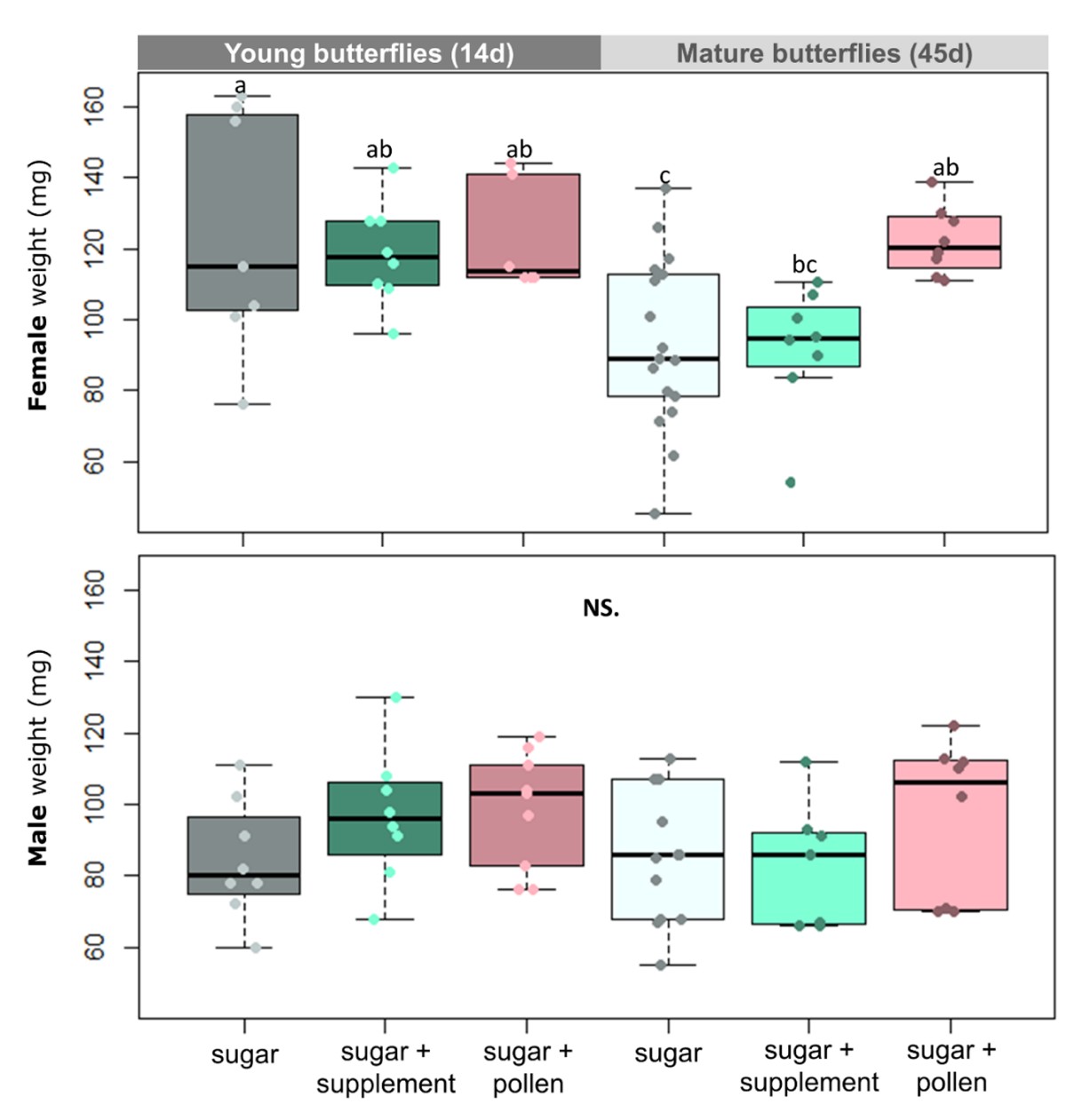
Figure 1 - Effect of diet and age on the fresh weight of females (top) and males (bottom) of H. erato. Butterflies were fed either sugar or sugar + supplement (Critical Care Formula) or sugar + pollen (from Lantana flowers). Young butterflies were collected after 14d of trial while mature butterflies after 45d. Legend: Different letters over the boxplots correspond to statistically significant differences (Two-ways ANOVA, Tukey HSD). NS = not statistically significant (p> 0.05). Lines in the middle of boxplots correspond to the median and boxes to the lower and upper quartile. Dots correspond to values of each analysed replicate/individual butterfly.
Table 1 - Effect of diet, age and sex on weight (grams per individual) of H. erato butterflies. The variables that have a significant effect on butterfly weight are marked in bold, with a * near their p value (p > 0.05). Diet treatments: sugar only, sugar + supplement, and sugar + pollen. Sex: female and male. Age: young (14d) and mature(45d).
| Variables | Three-way ANOVA |
|---|---|
| Diet | F2-95= 4.597, p= 0.012* |
| Age | F1-95= 7.287, p= 0.008* |
| Sex | F1-95= 25.055, p= 2.5 x 10-6* |
| Diet:Age | F2-95= 2.016, p= 0.139 |
| Diet:Sex | F2-95= 0.486, p= 0.617 |
| Age:Sex | F1-95= 4.520, p= 0.036* |
| Diet:Age:Sex: | F2-95= 1.791, p= 0.172 |
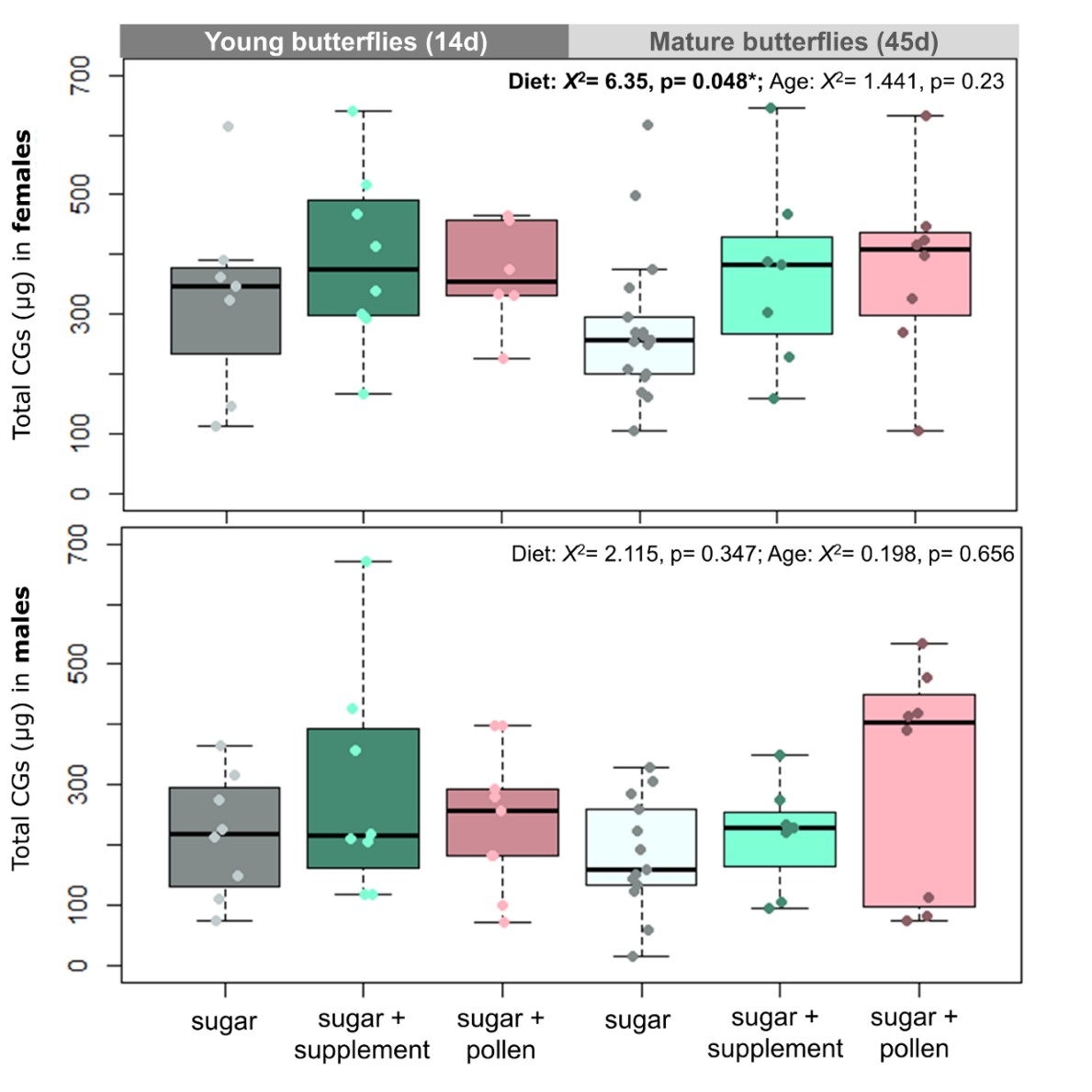
Figure 2 - Effect of diet and age on cyanogenic glucosides content of females (top) and males (bottom) of H. erato. Butterflies were fed either sugar or sugar + supplement (Critical Care Formula) or sugar + pollen (from Lantana flowers). Young butterflies were collected after 14d of trial while mature butterflies after 45d. Statistical analyses on the top of the plots correspond to Kruskal-Willis on Diet and Age for the subsets. Lines in the middle of boxplots correspond to the median and boxes to the lower and upper quartile. Dots correspond to values of each analysed replicate/individual butterfly.
Access to pollen only affects the chemical defences of females
Males and females increased their CG content after eclosion (Figure S2) and kept their defences through adulthood, indicating that they intensively biosynthesize these compounds. Curiously, diet only affected the CG content of females (Figure 2. Kruaskal-Wallis, Females: X2= 6.35, p= 0.048*; Males: X2= 2.115, p= 0.347), with butterflies having access to amino acid (supplement or pollen) showing greater CG content than those fed sugar alone. Young and mature butterflies of both sex had similar CG content (Figure 2. Kruaskal-Wallis, Females: X2= 1.441, p= 0.23; Males: X2= 0.198, p= 0.656).
Access to pollen delays reproductive senescence
Adult diet affected egg laying in mature butterflies of H. erato (Figure 3. Kruskal-Wallis, X2= 0.569, p= 0.017*), but not in young ones (Figure 3. Kruskal-Wallis, X2= 0.569, p= 0.752). Young females (14d) laid similar number of eggs regardless of their diet. In contrast, mature females (45d) that had access to pollen laid more eggs than butterflies that had access to sugar only and sugar + supplement. This indicates that access to pollen delays reproductive senescence in Heliconius.
In contrast to our expectations, adult diet did not affect parental allocation in the chemical defences (CG) of their eggs. Eggs of young (ANOVA, F2-10= 0.56, p= 0.588) and mature butterflies (ANOVA, F2-17= 0.09, p= 0.914) had similar concentrations of CG regardless of the diet of their parents.
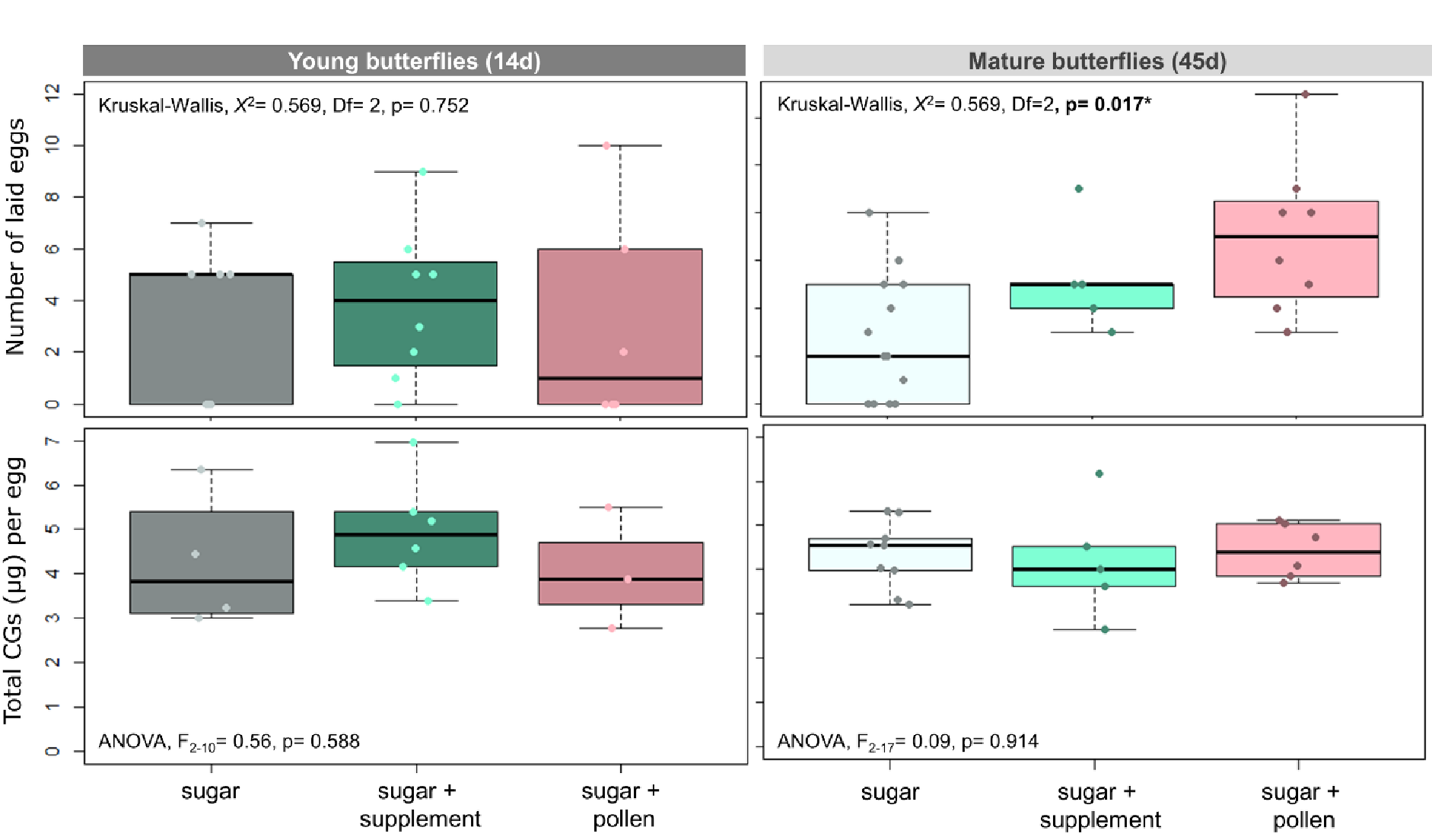
Figure 3 - Number of laid eggs per female during fertility test (top) and cyanogenic glucose content per egg (bottom). Butterflies were fed either sugar or sugar + supplement (Critical Care Formula) or sugar + pollen (from Lantana flowers). Young butterflies were collected after 14d of trial while mature butterflies after 45d. Lines in the middle of boxplots correspond to the median and boxes to the lower and upper quartile. Dots correspond to values of each analysed replicate/total eggs laid by each butterfly.
The nutritional uniqueness of pollen
Although the CCF supplement had far more protein (651.70 ± 19.97 µg per mg DW) than the exudate of Lantana flowers (1.71 ± 0.45 µg per mg DW) (Table S3), it did l not lead to better improvements in the butterfly fitness traits. Thus, pollen might have an amino acid profile that fits better the nutritional needs of Heliconius.
Discussion
When does access to pollen start to be important and for whom?
Pollen-feeding eases energetic contains and allows Heliconius to optimize multiple life-history traits simultaneously, aiding the maintenance of fertility, body weight and chemical defences during their prolonged adult-lifespan (Fig. 1, 2 and 3), as we hypothesized. Nevertheless, we found agespecific responses to this dietary novelty, as access to pollen has an effect on old butterflies (45d) of H. erato, but not on the young ones (14d). This is the first time to our knowledge that the effect of pollen-feeding on multiple life-history traits has been evaluated controlling for age. Studies supplementing the diets of other nectar-feeding long-lived nymphalids (Polygonia c-album, Maniola jurtina) with amino acids have not found an improvement in life-history traits (Karlsson & Wickman, 1989; Grill et al., 2013). Our results therefore emphasize that adaptations were required to make use of pollen-derived amino acids in Heliconius (Dunlap-Pianka et al., 1977; Boggs et al., 1981)
The age-effects also reveal that the balance between larval and adult derived resources changes over the life course. It is possible that the physiology of young Heliconius butterflies, including their fertility, initially relies mostly on resources acquired during larval feeding, as in the vast majority of butterflies and moths. Nevertheless, as Heliconius butterflies live relatively very long adult lives, the reservoir of larval derived resources likely becomes depleted over time, such that the presence of nitrogen in their adult diet becomes a crucial factor for the maintenance of the homeostasis. This is consistent with field data showing that older Heliconius butterflies generally collect more pollen then the young ones (Boggs et al., 1981), which may indicate greater motivation as larval resources deplete.
This implies that studies evaluating the importance of pollen-feeding during adulthood for Heliconius butterflies likely need to be performed for periods longer than a month. Cardoso & Gilbert (2013) did not observed differences between the cyanide concentration from 20d old Heliconius butterflies (H. ethila, H. hecale and H. charithonia) fed only sugar and supplemented with amino acids, as we observed here for females. The authors discussed the importance of larval diet shaping the chemical defences of young Heliconius butterflies. Additionally, the pheromone bouquet of 14 day old Heliconius males (genital and androconia) was also not affected by access to pollen during adulthood, only by the hostplant species used during larval feeding (Darragh et al., 2019). Indeed, the results seen here would probably be more striking if we have maintained the experiment for more than 45 days. Combined, these studies emphasize that the importance of the resources accumulated during larval feeding for young butterflies and the latter importance of pollen during adulthood.
It is a common knowledge among researchers breeding Heliconius under insectary conditions that they die sooner without access to pollen/amino acids in their adult diet. In this study, 45 days was not enough to observe differences in mortality between diet treatments (1-3 butterflies in all treatments, mostly whiting first week of eclosion), contrary to previous findings (Dunlap-Pianka et al., 1977). As previous experiments used H. charithonia ((Dunlap-Pianka et al., 1977), while we used H. erato, this could indicate that different Heliconius species might rely on the amino acids acquired during adult-feeding sooner than others. Nevertheless, the previous work had few replicates (N=8 in total, N=3 after 40 days.) and might have underestimated how long H. charithonia can live on average without pollen. How different Heliconius species respond to the lack of pollen is an interesting question for the future.
Do females pay a higher cost for reproduction than males when resources as scarce?
This study demonstrates that access to nitrogen delays reproductive senescence in H. erato females, as old females (45 d) supplemented with pollen lay as many eggs as young females (14d), whereas females fed only sugar lose fertility throughout adulthood (Fig. 3). This corroborates with the earlier works (Dunlap-Pianka et al., 1977) demonstrating that H. charithonia that have access to pollen can keep daily egg-laying rates until their natural death (up to 72d), however they continuously decrease their egg-production and reach ovarian depletion when pollen is absent.
The disparity between how females and males of the same species alter their life-history dynamics in response to resource availability has intrigued evolutionary ecologists. Without pollenfeeding, H. erato females lose weight (Fig. 1) and decrease their chemical defences (Fig. 2) as they get older, whereas males do not. As females collect significantly more pollen than males in the wild (Boggs et al., 1981), it could be that females feed more than males and therefore their fitness is more impacted by diet. Regardless, only females of H. erato were strongly affected by adult diet and this was reflected in their fertility, which might suggest that females are paying a higher energetic cost for reproduction than males when access to amino acids is limited. Even though diet did not affect male weight or CG content, we cannot discard the possibility that the effect of diet on fertility might be associated with other male fitness traits, such as sperm viability and quality of nuptial gifts.
Some of the old female butterflies in the cage supplemented with pollen had a strong smell of anti-aphrodisiac (personal observations) suggesting that they recently re-mated. Although remating was not expected in this experiment, since H. erato belongs to the monoandrous clade of Heliconius (Beltrán et al., 2007) and rarely re-mates in the wild (Cardoso et al. 2009; Walters et al., 2012), the insectary conditions might have induced them to re-mate. Re-mating would allow the transference of more nuptial gifts, which includes CGs, from the male to the female (Cardoso & Silva, 2015), diluting the effect of pollen supplementation on male chemical defences and body weight. Further studies of spermatophore quality will be necessary to unravel the effect of pollenfeeding on the fitness of Heliconius males.
Do high condition adults lay better protected eggs?
Many insects protect their eggs by transferring defensive compounds to them, which can improve offspring establishment. Thus, we hypothesized that butterflies with access to pollen would produce eggs with more CGs, as these compounds are not toxic when intact and can be stored in high concentrations. Old females of H. erato that had access to pollen are heavier (Fig. 1), more toxic (Fig. 2) and laid more eggs (Fig. 3) than old females that had access to sugar only. Contrary to our predictions, this does not translate into a higher investment in the chemical defences of their offspring (Fig. 3). Eggs of butterflies from all ages and diets have similar CG content, which suggests that this process is tightly regulated - butterflies might lay fewer or more eggs depending on their diet, but all eggs have a similar level of chemical defences. The amount of CG per egg observed here is similar to other heliconiines (Nahrstedt & Davis,1983; 1985; Pinheiro de Castro et al., 2020).
Our data demonstrate how strongly H. erato biosynthesize CG during adulthood to maintain their defences, while also investing in the protection of their offspring, corroborating previous findings (Pinheiro de Castro et al. 2020; Mattila et al., 2022). Considering that a Heliconius female can live for 45 days, lays ca. 10 eggs per day (Dunlap-Pianka et al., 1977) and each egg has on average 3 µg of CG (Fig. 3), egg-laying would therefore result in a depletion of over 1000 µg of CG from a female butterfly, which can be more than their whole reservoir of chemical defences at any one time (Fig. 3). In contrast, male contributions for offspring chemical defences seems minimal (Cardoso & Gilbert, 2007). Pedigree experiments with H. erato also found strong maternal effects on offspring toxicity, but no paternal effects (Mattila et al., 2021).
Mattila et al. (2022) demonstrated that Heliconius butterflies keep their CG concentration at high levels during adulthood until their natural death. Indeed, if these aposematic butterflies lost their toxicity as they age, this would dilute the protection signal of their colour pattern. Thus, there is probably strong selection for Heliconius to maintain toxicity as they age, but it is likely challenging to maintain these levels while reproduction depletes their chemical reservoir (Fig. 2 and 3).
Moreover, valine and isoleucine are used as substrate for the biosynthesis of aliphatic CGs (Nahrstedt & Davis, 1983). These are essential amino acids that have to be acquired by diet (not produced by animals) and they tend to be abundant in pollen (Gilbert, 1972). This suggests a strong effect of pollen-feeding on chemical defences in Heliconius. Yet, a lack of pollen/amino acids during adult-feeding does not affect the chemical defences of young Heliconius butterflies. As already discussed, access to pollen would become crucial at later stages of adulthood, but the remaining question is: where did the valine and isoleucine used for CG biosynthesis come from during the first weeks of Heliconius adulthood in the control group (sugar only)? A recent comparative genomic study has found that two hexamerins, storage proteins, have been duplicated multiple times in heliconiines (Cicconardi et al., 2023). Hexamerins might provide valine and isoleucine for CG biosynthesis during the beginning of their adulthood, if pollen is not available. Moreover, valine and isoleucine might be produced by bacteria in the microbiome of Heliconius butterflies, as happens for other insects (Jing et al., 2020), a hypothesis that can be investigated in the future.
Conclusion
In summary, although the link between pollen-feeding, fertility and chemical defences in Heliconius butterflies is clear, these interactions are more complex than initially predicted. We demonstrated that there is an age-specific and sex-specific effect of pollen-feeding on life-history traits. Older females supplemented with pollen were heavier, more toxic and laid more eggs than those in the control diets, suggesting that this dietary innovation has eased energetic constraints and led to optimization of multiple life-history traits.
Acknowledgments
Preprint version 5 of this article has been peer-reviewed and recommended by Peer Community In Evolutionary Biology (https://doi.org/10.24072/pci.evolbiol.100648; Briscoe, 2023). We thank Adriana Briscoe, Carol Boggs and Caroline Muller for the excellent revision of this manuscript. We thank the Lepidoptera international community for all the valuable discussions on life-history evolution. We thank the Zoology Derpatment of Cambridge University for hosting this project, especilly all the technical support staff that enabled our reserach activities. We also thank the staff at the PLBIO-PLEN-KU, for the assitance in the target-metabolomic analyses, especially Mariela Alejandra González Ramírez for the maintenance of the analytical instruments and Mohammed Saddik Motawie for the chemical standars used in these analyses. EC and CJ thank the Horizon 2020 - Merie Curie Actions and UKRI for the finantial support in this project. AM thank the Research Council of Finland for her fellowship. All authors of this manuscript declare no conflict of interest.
Funding
The researchers involved in this project were supported by the following grants: H2020 Marie Skłodowska-Curie Actions (841230), ERC Starter Grant (758508), Biotechnology and Biological Sciences Research Council (BB/R007500), Academy of Finland (Grant no. 286814), UKRI-NERC (NE/N014936/1), UKRI-NERC (NE/W005131/1).
Competing interests
The authors declare that they have no competing interests.
Data, script, code, and supplementary information availability
The metadata, coding script and raw chemical data associated with this publication are available in the Apollo repository from University of Cambridge at https://doi.org/10.17863/CAM.92867 (Pinheiro de Castro et al. 2023).
Supplementary material
Supplementary Methods
Sample preparation and protein quantification
Pollen and nectar samples of Lantana sp and Psiguria sp were collected by pipetting and resuspending multiple stime ultra-pure water into their flowers (10 µL per flower). Three pooled replicates were collected per species, each composed of the exudate of 10 flowers for Psiguria and 20 flowers for Lantana. For the supplement, 20 mg Critical Care Formula (CCF) were mixed in 10 mL ultra-pure water per replicate. Protein assay was performed using the PierceTM660 nm method (ThermoFisher) following the manufacturer instruction: 100 µL of each sample was incubated into 1.5 mL of Pierce Reagent and their absorbances read at 660 nm against a blank (100 µL ultra-pure water and 1.5 mL Pierce Reagent). A standard curve composed of Bovine Serum Albumin solutions at different concentrations was also assayed and the established regression equation used to calculate the amount of protein per sample. The remaining flower samples were dried in a SpeedVac (medium speeds) for 2 hours to establish their dry mass.
Supplementary Tables
Table S1. Normality test (Shapiro-Wilk) on the analysed variables. * correspond to variables that have a normal distribution (p > 0.05).
| Variables | Shapiro-Wilk |
|---|---|
| Butterfly weight (mg) | W= 0.987, p= 0.306* |
| Laid eggs | W= 0.926, p= 0.0056 |
| Butterfly CG content (ng per butterfly) | W= 0.946, p= 0.0001 |
| Egg CG content (ng per egg) | W= 0.972, p= 0.668* |
Table S2. Homogeneity of the variances (Levene’s Test) on the analysed variables, controlling by diet, age and sex. * Correspond to variables where the homogeneity assumption is met (p > 0.05).
| Variables | Levene’s Test |
|---|---|
| Butterfly weight (mg) | F11-95= 1.468, p= 0.157* |
| Laid eggs | F5-41= 0.363, p= 0.871* |
| Butterfly CG content (ng per butterfly) | F11-94= 0.628, p= 0.801* |
| Egg CG content (ng per egg) | F5-27= 0.636 p= 0.674* |
Amino Acid Source | Protein Concentration (µg/mg DW) |
|---|---|
CCF supplement | 651.70 ± 19.97 |
Lantana sp pollen and nectar extract | 1.71 ± 0.45 |
Psiguria sp pollen and nectar extract | 9.20 ± 0.97 |
Table S3. Protein concentration (µg/mg DW) in supplement and flowers extracts used as a amino acid source to feed Heliconius butterflies.
Supplementary Figures
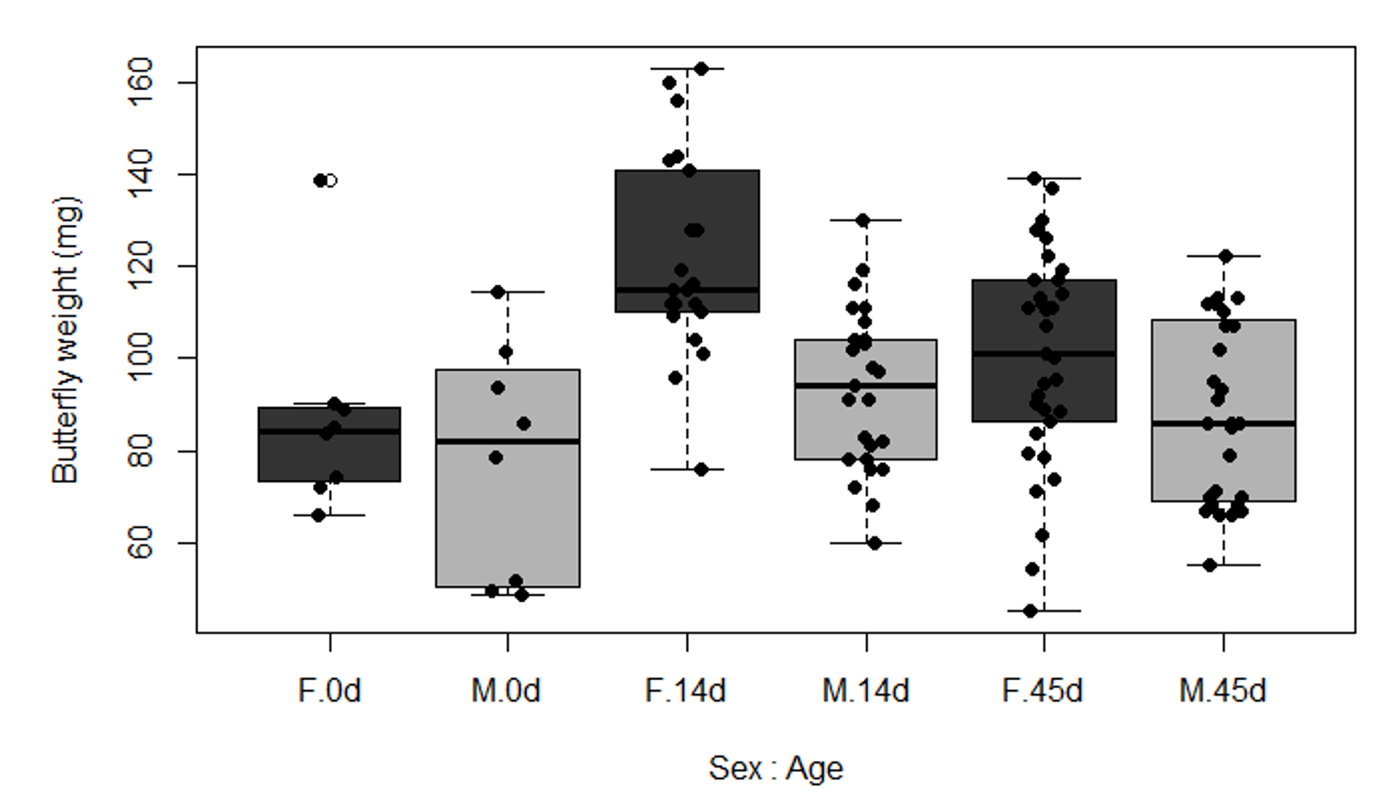
Figure S1. Butterfly weight of H. erato females (F) and males (M) at different ages (0d, 14d and 45d after eclosion).
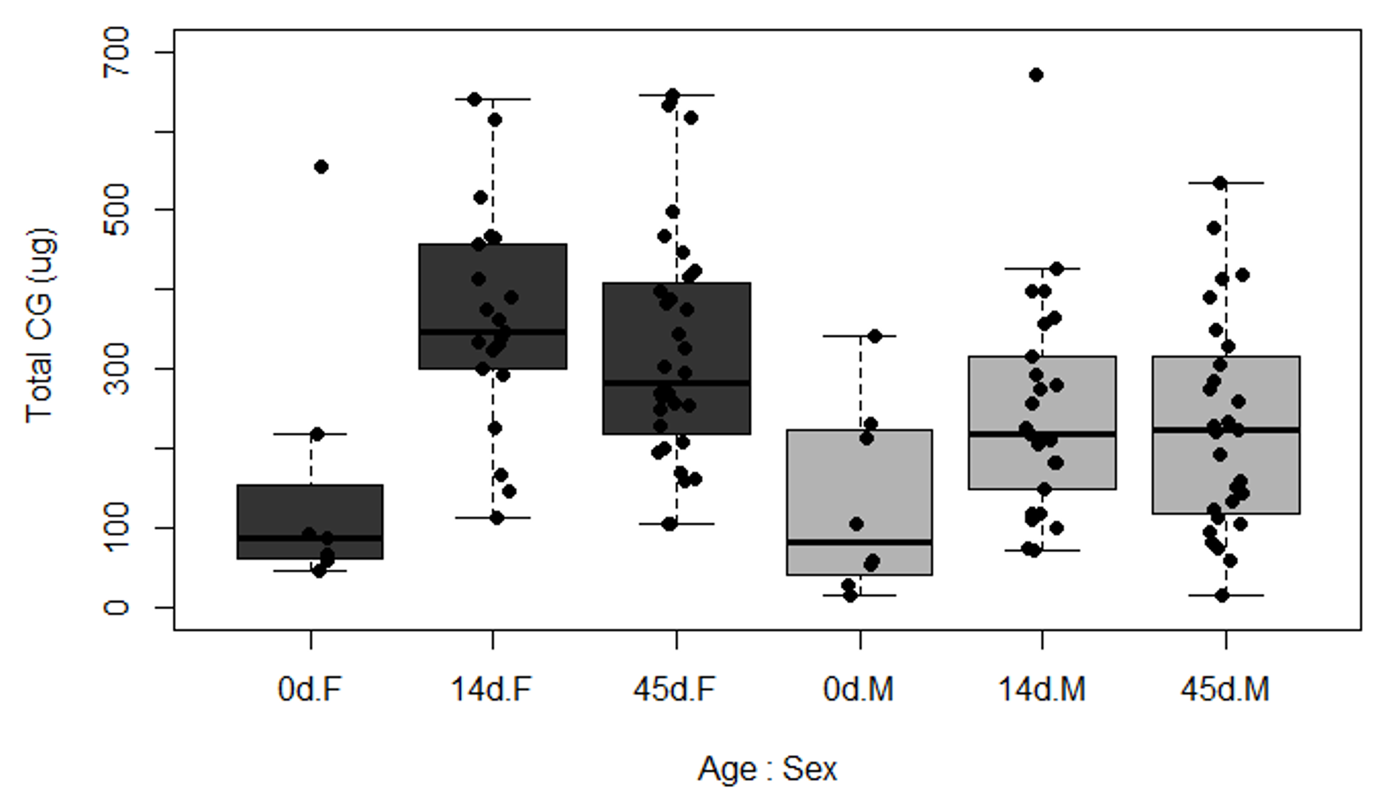
Figure S2. Total content of cyanogenic glucosides (CG) in females (F) and males (M) of H. erato at different ages (0d, 14d, and 45d after eclosion)
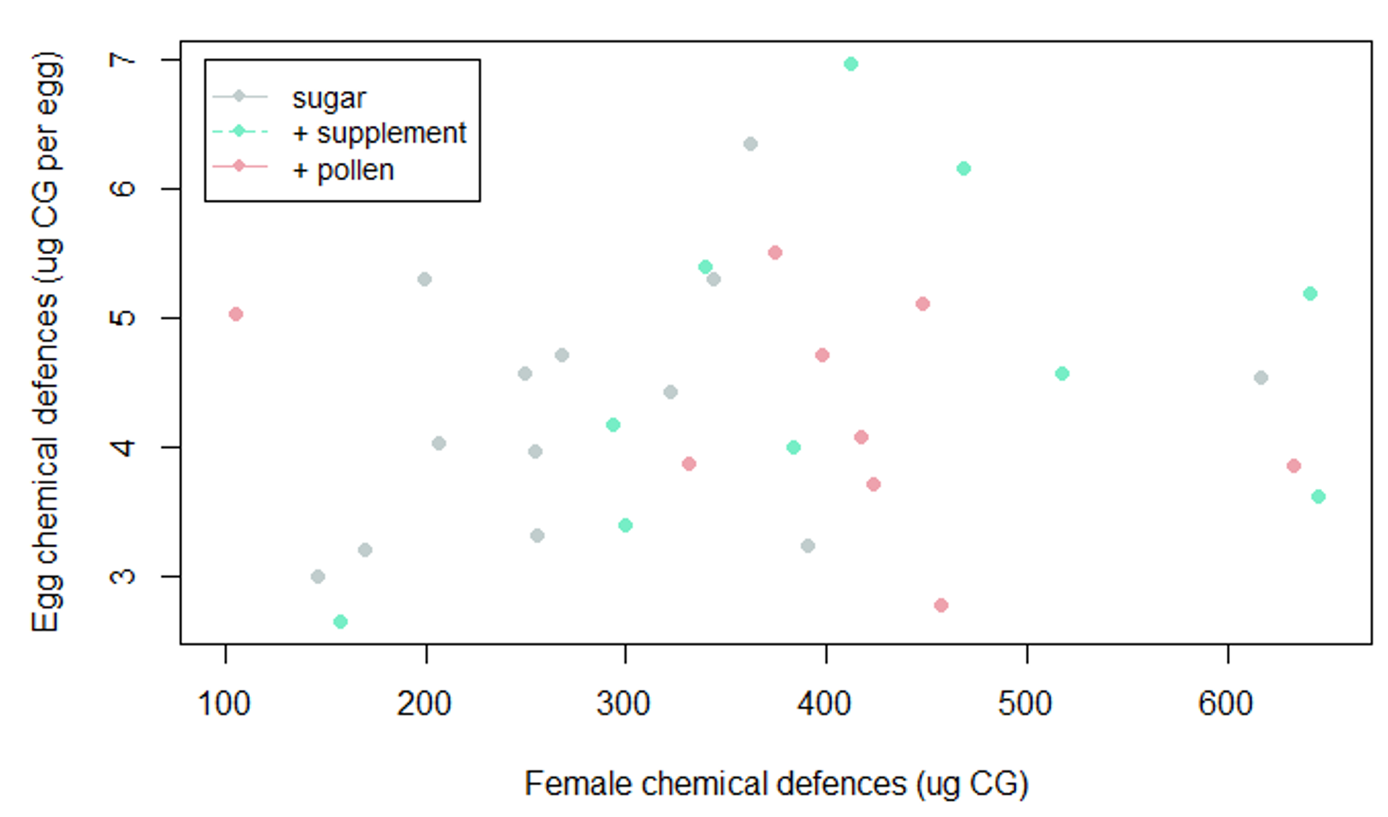
Figure S3. Relation between egg chemical defences and female chemical defences. The correlation is not significant (R2= 0.033, F1-30= 1.021, p= 0.32).