
 CC-BY 4.0
CC-BY 4.0
Introduction
Understanding the factors that promote the evolution of trait diversity is a major goal of ecology and evolutionary biology. Animal signals are particularly noteworthy examples of highly diverse traits in the natural world. They are ubiquitous, occurring across all sensory channels, from acoustic to chemical (Baeckens et al., 2018; Gallagher et al., 2024; Ord & Martins, 2006), and serve vital functions for individuals, such as species and individual recognition, sexual signaling, and predator defense (Laidre & Johnstone, 2013). While signals are found in most species, some species exhibit much greater signal diversity than others (Schaefer & Ruxton, 2015). Profound interspecies differences in signaling diversity are evident across all taxa, including mammals, reptiles, birds, and insects (D’Ammando & Bro-Jørgensen, 2024; Miles et al., 2020; Nehring & Steiger, 2018; Ord et al., 2001). For example, some species of birds, such as the spot-breasted oriole (Icterus pectoralis), produce gestural display with low signal diversity while others, like the shiny cowbird (Molothrus bonariensis), exhibit a high diversity, with a wide range of variations (Miles et al., 2017).
Signal diversity is typically studied by quantifying the number of unique elements expressed in a signal. These unique elements can be specific to a single species or shared among multiple species, and assessing the similarity between elements allows to evaluate how these elements are distributed across the phylogeny (Price & Lanyon, 2002). Similarities in signals have been mainly studied in the context of mimicry (Raguso, 2008), but they also occur in other evolutionary contexts that require further investigation to understand why some species share certain elements while others do not. Similarities in the way emitters produce signals and receivers perceive them are expected to explain similarities in the signals themselves. For instance, visual modeling of predators has revealed cases where insects share warning signals (Penacchio et al., 2024), and specific chemical compounds may be more effective in attracting pollinators (Gervasi & Schiestl, 2017). The similarities in signal function may therefore explain why some signals share elements or physical characteristics but this has been poorly studied (Delhey et al. 2023).
Acoustic signals are highly variable between species and thus serve as good models to tackle questions of evolution towards signal diversification or similarity in signal composition at the interspecific level. They are widely used in terrestrial vertebrates, notably in the oscine passerines, which stand out as one of the rare animal groups where individuals acquire their vocal signals through learning (Tyack, 2019). In this group, songs are acoustic signals composed of different sound units, produced by a sophisticated vocal organ, controlled by a neurobiological system, and involved in reproductive functions (however, see Rose et al. (2022)). Songs are composed of several acoustic elements defined by distinct physical time and frequency parameters named syllables. The diversity of song, also called song complexity or song elaboration, is measured by the within or between song diversity (syllable or song repertoire size) or the syllable versatility (syllable diversity per unit of time). Song composition, on the other hand, relies on the identification of discrete categories based on the sound characteristics of the acoustic units that make up a song. These acoustic units (or syllables) are grouped into categories according to their acoustic properties (Kershenbaum et al., 2016). For instance, in their study, Price and Lanyon (2002) define different categories based on specific acoustic criteria, such as trills, clicks, rattles, or whistles. The authors then explore how these sound categories are distributed across the phylogeny. Often, it is assumed that the observed acoustic properties stem from different sound production mechanisms used by the signallers. This approach enables comparisons across species by identifying shared or distinct acoustic features within a given taxonomic group (Odom et al., 2021; Sung et al., 2005).
Sexual selection is one of the main factors acting on the evolution of animal signals’ (Darwin, 1871) diversity (Schaefer & Ruxton, 2015) and composition (Schwark et al. 2022). In birds, the role of sexual selection on the evolution of song diversity has been extensively documented in both wild and laboratory studies (Byers & Kroodsma, 2009). The ability to produce a large repertoire correlates with cognitive ability (Boogert et al., 2008), body condition (Kipper et al., 2006; Soma et al., 2006) and memory capacities which can be impaired by developmental stress (Nowicki et al., 2000; Spencer et al., 2003; Zann & Cash, 2008), and hence large repertoires are thought to correlate with individual quality and be under sexual selection. However, numerous counterexamples also exist (Soma & Garamszegi, 2011; Garamszegi & Møller, 2004), suggesting that diversity does not always evolve under sexual selection (Robinson & Creanza, 2019; Gil & Gahr, 2002). At the interspecific level, few studies have investigated the relationship between song diversity and sexual selection (Snyder & Creanza, 2019), with recent comparative analyses revealing mixed results. For instance, one study found no significant link between mating systems and syllable repertoire size (Snyder & Creanza, 2019), while another demonstrated a positive correlation between intra-song diversity and the frequency of extra-pair paternity (Hill et al., 2017a). Regarding song composition, an effect of sexual selection can be expected at the interspecific level as some song elements potentially serve as an indicator of mate quality and may be more present under strong sexual selection. For example, some song elements are more challenging in terms of biomechanics due to their requirement for precise motor control (Suthers et al., 1999; dos Santos et al., 2023; Goller, 2022). This is well illustrated by canaries that produce large two-voice notes syllables at high rates which are controlled independently by each brain hemisphere, making it difficult to produce quickly and the individuals that do it are preferred by potential mates (Suthers et al., 2012). Another example is the buzz-like element in the song of water pipits (Anthus spinoletta), which has been shown to predict pairing success (Rehsteiner et al., 1998).
Sociality appears to be another driver of signal diversity (Roberts & Roberts, 2020; Ord & Garcia-Porta, 2012; Peckre et al., 2019) which could also act on song composition (Morinay et al. 2013). Under the ‘social complexity hypothesis for communication’, more diverse and complex social interactions select for greater signal diversity (Freeberg et al., 2012; Peckre et al., 2019). For acoustic signals, this hypothesis has been supported in a wide range of animals, mostly mammals, like rodents (Blumstein & Armitage, 1997; Pollard & Blumstein, 2012) and primates (Fichtel & Kappeler, 2022; McComb & Semple, 2005), but also in birds (Krams et al., 2012, Leighton & Birmingham, 2021). In mammals, group size, which is expected to increase social complexity, has been shown to impact signal diversity at the interspecific level (Pollard & Blumstein, 2012). In birds, only one study on the Carolina Chickadee (Poecile carolinensis) has experimentally tested the effect of group size on acoustic diversity, showing that birds in larger groups used calls with greater diversity of note types and note combinations (Freeberg, 2006). To date, however, no study has examined the association between colony size and song diversity at the interspecific level in birds. The ability to produce a large song repertoire size requires the ability to (i) biomechanically produce significant acoustic variation and (ii) possess extended memory capacities to memorize those variations. These physical and neurobiological capacities may have been selected within a social communication context. For song composition, there is restricted evidence for an effect of sociality. However, we can hypothesize that some song acoustic features may allow for finer individual identification in larger colonies and/or might be more efficiently transmitted than others among the background noise at the colony because they maximize the signal-to-noise ratio (Aubin & Jouventin, 1998).
In addition to the influence of sexual selection and sociality, the diversity of birdsong is also expected to be constrained by phylogeny. Examining the interplay between phylogenetic history and the evolution of birdsong, for instance by measuring the phylogenetic signal, the degree to which species' trait similarities reflect their evolutionary relationships, allows for the interpretation of evolutionary patterns. Evidence of a phylogenetic signal in vocalisations has been found in several bird families (Medina‐García et al., 2015; Mejías et al., 2020; Päckert et al., 2003; Rivera et al., 2023). With respect to song diversity, studies have reported both low (Crouch & Mason‐Gamer, 2019) and high (Tietze et al., 2015; Snyder & Creanza, 2019) levels of phylogenetic signal. Regarding song composition, closely related species may share more acoustic features of their syllables due to shared bio-mechanisms of producing vocalisations or to ecology. Price and Lanyon (2002) investigated homology in various song parameters within a clade of 12 species of Oropendolas and found high conservatism in many features, suggesting a strong genetic control of song in this bird family (see also Sung et al., 2005).
Weaverbirds (Ploceidae) are an interesting study system to test the effects of sexual selection, social selection and phylogenetic constraints on song diversity and composition at the interspecific level. They exhibit diverse social mating systems, from polygamy to monogamy (del Hoyo et al., 2010) and variation in mating system has been demonstrated to be generally associated with variation in the intensity of sexual selection across species (Shuster, 2009). Additionally, they have variable social systems, from solitary to colonial, with colony sizes ranging from a few individuals to several thousand (del Hoyo et al., 2010; Oschadleus, 2020). Using comparative analysis, we investigated the association between song diversity and composition and (i) social mating system, which is used as a proxy for sexual selection and (ii) colony size, used as a proxy for sociality. Moreover, we considered phylogenetic similarity between species to assess the effect of shared history on these two song variables. Our predictions for mating system, colony size and phylogeny are summarized in Table 1. We controlled for habitat openness as weavers occupy diverse habitats, including woodlands, tropical forests, grasslands, wetlands, and savannahs (del Hoyo et al., 2010). Due to physical constraints imposed by the medium of transmission on the sound propagation, habitats might have variable effects on song (Morton 1975, but see Freitas et al., 2025) and both positive and negative effects have been found on song diversity (Cicero et al., 2020; Crouch & Mason-Gamer, 2019; Hill et al., 2017b; Leighton & Birmingham, 2021). We measured song diversity and acoustic composition of respectively 95 and 60 species of weaverbirds. Song diversity is represented by syllable repertoire size (also called syllable diversity, within-song diversity or intra-song repertoire size in other studies) and is calculated as the number of different syllables per song. Acoustic composition corresponds to the presence/absence of syllable elements which are defined based on several acoustic criteria describing the general shape of each syllable.
Table 1 - Hypotheses, rationale and predictions of the study.
Variable | Hypothesis | Rationale | Prediction |
|---|---|---|---|
Sexual selection increases song diversity | Song diversity signals individual condition | Polygamous species exhibit a larger syllable repertoire size than monogamous species | |
Song diversity | Sociality increases song diversity | Richness in elements allows for more precise information | Species living in larger colonies exhibit a larger syllable repertoire than species living in smaller ones |
Phylogenetic history affects song diversity | Relatives share common traits | Variation in syllable repertoire size parallels phylogenetic relationships | |
Sexual selection affects song composition | The acoustic characteristics of certain elements are biomechanically more difficult to produce | More similarity in the types of syllables produced within polygamous species, as well as within monogamous species | |
Song composition | Sociality affects song composition | The acoustic characteristics of certain elements allow to code more precisely for individuality or favor transmission in noisier groups | More similarity in the types of syllables produced within species living in larger colonies, as well as within those living in smaller colonies |
Phylogenetic history affects song composition | Relatives share common traits | Species with a longer shared history produce types of syllables that are closer to one each other than those having diverged a longer time ago |
Material and methods
Data collection
The song data were collected from online sound archives Xeno-Canto (53% of the sampled individuals), Macaulay (24%), the British library (9%), CD storage media (Chappuis, 2000) (6%) and from recordings performed by us in the field in South Africa in January 2021 and in Sao Tomé in June 2022 (8%). Recordings in the field were made using a Sennheiser microphone (MKH70 with K6 power module) connected to a Marantz PMD661MK2 digital recorder, on focal individuals. Recordings targeted males performing courtship in presence of females.
We assessed the sound quality of all the audio tracks by listening and through visual inspection of sound spectrograms. We paid attention that in the files only a single individual was singing and that the song clearly emerged from the noise. Only .WAV files, an audio format not compressing high frequencies, with a sampling rate of 44.1 kHz, were analysed.
In total, we analysed 3577 songs of 365 individuals (mean number of songs per individual is 3.71, sd = 4.66) from 95 species (mean number of individuals per species is 3.84, sd = 1.97).
Songs, syllables and category labeling
Recording files were analysed using Avisoft SASLab Pro v.4.3.01. For each recording we first identified the songs, by defining a song as a sequence of at least 2 different sound units separated by less than 1.5s of silence. We then identified and labeled syllables in each song (see detailed method in supplementary material). A syllable was defined as a sound unit represented by a continuous trace on the spectrogram not interrupted by more than 0.015s (see Figure S1 for an example of song and syllable labeling).
To analyse song composition, we defined categories of syllables according to several distinct general acoustic properties such as the presence of harmonics, the characteristics of the frequency modulation, the presence of pulse sounds, the presence of two-voice phenomenon (see Table S1 for a detailed description of syllables categories), and then attributed a category to each identified syllable based on its temporal and spectral properties. We identified 59 categories. While syllables are defined for each new song, categories are common for all species and therefore allow between species comparison of song composition.
Response variables
Song diversity
We estimated the syllable repertoire size (hereafter song diversity) for each song by counting the number of different syllables contained in the song. Because several recordings contained multiple songs per individual, song diversity for each individual was calculated as the mean for all the produced songs by a given individual. Song diversity was estimated in all species (95 species) retaining individual values without averaging them at the species level.
Song Composition
We estimated the composition of songs for each species by integrating the 59 syllable categories into a presence-absence matrix. For each species, we assessed the presence or absence of each category in the entire set of songs recorded for that species (combining all recordings and individuals). A category is considered present in a species if, among all the syllables recorded for that species, at least one syllable belongs to that category.
The presence-absence matrix of syllable categories at the species level was then transformed into a distance matrix to be included as a response variable in our model. For each species pair, we calculated the Jaccard distance, a measure of similarity between two sets of elements (resulting in 1770 combinations), and this distance was subtracted from 1 to transform it into an index of similarity to ensure a better interpretation of the model. These interspecies similarities represent the response variable in our model.
For this variable, only species with comprehensive sampling were included in the analysis. To estimate sampling effort, accumulation curves were generated for each species (Figure S2). Only the species in which the repertoire curve reached a plateau were included (60 species out of the 95). Even though our sample size is adequate, since this measurement is qualitative and based on the presence/absence of features, it may be more sensitive to sampling effort than the song diversity measure. Accumulation curves could not be generated for song diversity, as syllables are redefined for each song.
Fixed predictor variables
We used mating system as proxy for sexual selection intensity. Mating system was classified as ‘monogamy’ or ‘polygamy’ (del Hoyo et al., 2010; Song et al., 2022) with polygamous species considered to be more likely to be under stronger sexual selection than monogamous species (Shuster, 2009). Colony size was used as a proxy for sociality and scored as an ordinal categorical variable decomposed into four levels representing various degrees of colony size: solitary (1 to 2 individuals), small (2 to 5 individuals), medium (5 to 15 individuals) and large (more than 15 individuals) (Oschadleus, 2020; del Hoyo et al., 2010). Habitat openness, was classified as ‘closed’ (forest, woodland) or ‘open’ (grassland, wetland, savannah) (del Hoyo et al., 2010; Song et al., 2022; Mikula et al. 2021; Mejías et al., 2020).
Regarding the song diversity model, predictors were included in the model as stated above. However, for the acoustic composition model, since the response variable is a distance between a pair of species, the predictor levels were transformed to reflect the difference or similarity between the two species in each species pair considered. Thus, the mating system variable obtained contains the levels ‘comparable’ and ‘different’. For a species pair where both species have the same level ‘monogamy’ or ‘polygamy’, the assigned level is ‘comparable’. Conversely, if one species has ‘monogamy’ and the other ‘polygamy’, the assigned factor is ‘different’. The same applies to the habitat variable. For the colony size variable, we aimed to capture differences in ordination between different levels. We established four levels (‘comparable’, ‘different’, ‘very different’, ‘extremely different’) based on the discrepancy between the levels of the two species in the species pair considered. Thus, the difference in colony size between a pair of species increases with the number of steps separating the colony size levels of one species from the other. For example, if one species has the level ‘large’ and the other ‘medium’, the assigned factor is ‘different’ and if one species has the level ‘solitary’ and the other ‘medium’, the assigned factor is ‘very different’.
Statistical analyses
Linear mixed models
To test how song diversity varied with mating system, habitat openness and colony size, we performed a negative binomial phylogenetic generalized linear mixed model (Mdiv). Because we have several individuals for each species we included the species as random effect to take into account the within species variance. To account for phylogenetic relatedness, a phylogenetic covariance matrix derived from a recent phylogeny (De Silva et al., 2017) was also included as random effect. The model equation is detailed in supplementary material (Eq. S1). To choose the appropriate link function we compared negative binomial and Poisson errors distributions using leave-one-out cross-validation to assess model fit relying on the expected log predictive density (Vehtari et al. 2017).
To test how song composition varied with mating system, habitat openness, colony size and phylogenetic relatedness, we performed a phylogenetic generalized linear mixed model with a multiple membership random effect structure to account for multiple species pairs comparisons (Mcomp). In this model, the response variable is a measure of similarity between a pair of species (the complement of the Jaccard distance, measured using the presence-absence matrix of categories), and the predictors are categorical variables assessing how similar the pairs are in terms of mating system, habitat openness, and colony size. Phylogenetic relatedness, a measure of similarity between pairs of species (the complement of genetic distance), is usually included as a random factor. However, since the response variable here is also a distance measure between species, we included phylogenetic relatedness as a fixed factor. The model equation is detailed in the supplementary materials (Eq. S2).
Both models were generated using the ‘brms’ package in R with the weakly informative default priors (Bürkner, 2017). For each, we ran four chains for 10,000 iterations with a burn-in period of 3000, thinned every 5 iterations. Chain convergence and autocorrelation was diagnosed and posterior predictive checks were performed to ensure model validity and fit to the observed data.
Phylogenetic signal
Phylogenetic signal indicates how closely related species tend to share similar traits (Blomberg et al., 2003). Assessing the phylogenetic signal offers insights into the evolutionary patterns of specific traits. Traits with strong phylogenetic signal likely evolved gradually over time, following a Brownian motion model of evolution while traits lacking phylogenetic signal may exhibit extreme flexibility (Revell et al., 2008). To test the phylogenetics constraints on weaverbird song diversity we measured the phylogenetic signal (which is similar to broad-sense phylogenetic heritability). It is necessary to estimate the phylogenetic signal on the residuals of a model because the assumptions regarding phylogenetic non-independence concern the residual errors of the regression model, not the individual traits themselves (Symonds & Blomberg, 2014). To estimate the phylogenetic signal we estimated the variance components of model Mdiv and calculated the phylogenetic intraclass correlation (ICC) using the QGglmm::QGicc() function (de Villemereuil et al., 2016).
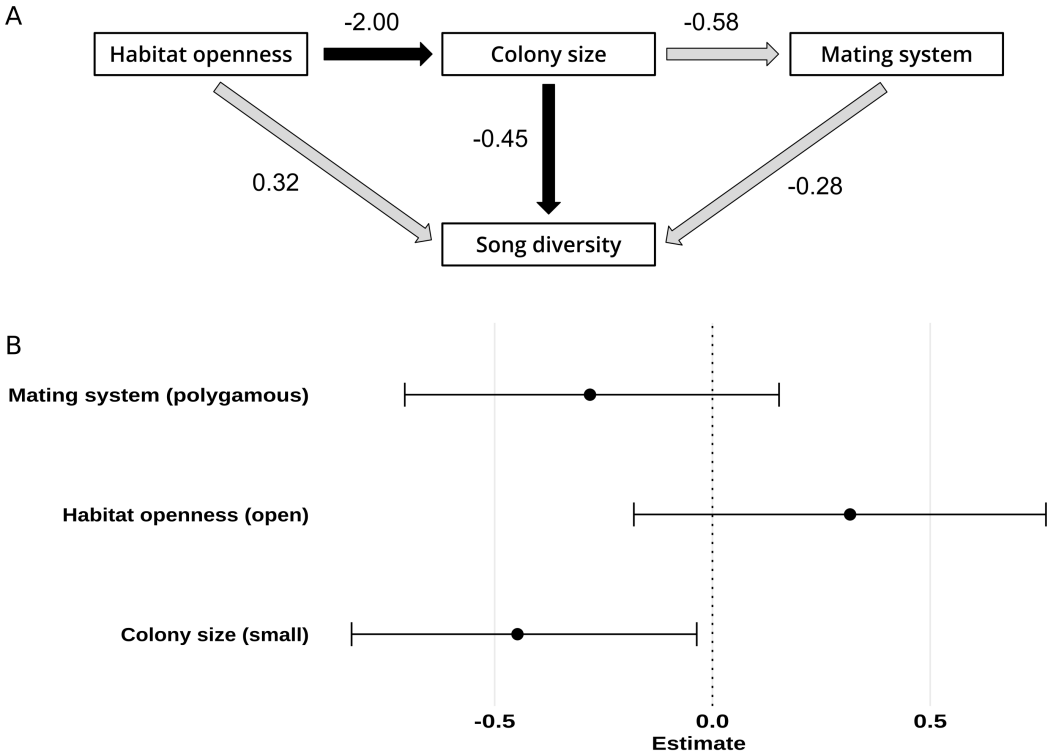
Figure 1 - Phylogenetic path analysis of the effect of ecology and life history traits on song diversity. (A) Directed acyclic graph representing the causal structure between the studied variables. Arrows indicate supposed direct causal effects, the strength of the effect is indicated by numeric values. Arrow colour indicates the significance of the effect: black indicates significant, and gray indicates non-significant. (B) Parameter estimates from the phylogenetic path model. Horizontal bars representing 95% confidence intervals obtained by bootstrapping and the points indicating the estimates. As in the bayesian phylogenetic multilevel model performed on song diversity (Mdiv), colony size effect is significant and habitat effect is non-significant. However, contrary to model Mdiv, mating system effect on song diversity is now non-significant.
Phylogenetic path analysis
In order to provide strong statistical support for the hypotheses tested, it is important that the measured variables are independent so parameters can be estimated precisely (Dormann et al., 2013). However, in our study, habitat openness, mating system, and colony size show some colinearity (see Figure S3). The covariation of these factors in weaverbirds has long been identified (Crook, 1964) and received further statistical support in a recent study (Song et al., 2022). In their study, Song et al. conducted a path analysis revealing that diet and habitat directly influence social foraging behavior that influences nesting behavior itself, which in turn influences the social mating system (for a broader overview, see Kvarnemo, 2018). These previous studies were therefore used to establish the hypothetical causal structure of mating system, colony size, habitat openness, and song diversity by constructing a directed acyclic graph (Figure 1A) (Arif & MacNeil, 2023; Wysocki et al., 2022). We then conducted a phylogenetic path analysis (PPA) to test these relationships (Hardenberg & Gonzalez-Voyer, 2013; Bijl, 2018). For this analysis we transformed the colony size variable into a binary variable merging ‘solitary’ with ‘small’ and ‘medium’ with ‘large’ to implement it in the PPA and log-transform the response variable to approximate normality. Also, contrary to Mdiv, in which we implemented one value of song diversity per individual in the model, for this analysis, we implemented one value of song diversity for each species (calculated as the mean value across individuals of the species). We used 500 bootstrap replicates to get the confidence interval estimates.
Results
Song diversity
Results from our model (Mdiv) revealed that the mating system and colony size are associated with song diversity, but habitat openness is not (Figure 2). The stronger effect size was for colony size. Larger colonies were associated with increased acoustic diversity in songs (Table S2). Solitary species exhibited lower mean diversity compared to those with large colony sizes (β = -0.66, 95%CI = [-1.00, -0.31]), as did species in small colonies (β = -0.28, 95%CI = [-0.56, -0.01]). By contrast, polygamous species, on average, exhibited lower song diversity than monogamous species (β = -0.29, 95%CI = [-0.56, -0.02]). Habitat openness, whether open or closed, did not appear to influence song diversity (β = 0.20, 95%CI = [-0.05, 0.50]). The phylogenetic signal of song diversity had a clear positive effect (median = 0.18, 95%CI = [0.05, 0.45]).

Figure 2 - Parameter estimates and posterior distributions from the bayesian phylogenetic multilevel model performed on song diversity (Mdiv). Estimates show a significant association of mating system and colony size with song diversity. For mating system, habitat and colony size, the references levels are respectively monogamous, closed and large. The posterior distribution of the estimates of each of the tested predictors are shown with horizontal bars representing 95% of the distribution and the points indicating the means.
Concerning the PPA (Figure 1B), the only variable having a clear effect on song diversity was colony size (β = -0.45, 95%CI = [-0.83, -0.04]). The model indicates that, all else being equal, species living in large colonies have, on average, a song diversity 36% higher than species living in small colonies. This result appears robust, as the effect size and its probability remain consistent across various comparisons. Indeed, whether comparing solitary species to colonial species, species living in large colonies to all other species, or when colony size is coded quantitatively, the effects remain similar. Neither mating system (β = -0.28, 95%CI = [-0.71, 0.15]) nor habitat openness (β = 0.32, 95%CI = [-0.18, 0.77]) clearly affected song diversity (Table S3).
Song composition
The results from our model (Mcomp) revealed that phylogenetic similarity and colony size had an effect on the acoustic composition of songs, but not mating system or habitat openness (Figure 3). According to our model, species that are closer in the phylogeny tended to produce songs with more similar acoustic composition (β = 0.034, 95%CI = [0.017, 0.051]). Furthermore, species with similar colony sizes exhibited more similar acoustic composition and species very different in their colony size (large colonies vs solitary species) had significantly different acoustic composition (β = -0.027, 95%CI = [-0.046, -0.009]). Differences in habitat openness, whether open or closed, did not affect the acoustic composition similarity (β = -0.007, 95%CI = [-0.016, 0.002]). Finally, differences related to the mating system also did not have an effect on acoustic composition similarity (β = 0.001, 95%CI = [-0.008, 0.010]) (Table S4).
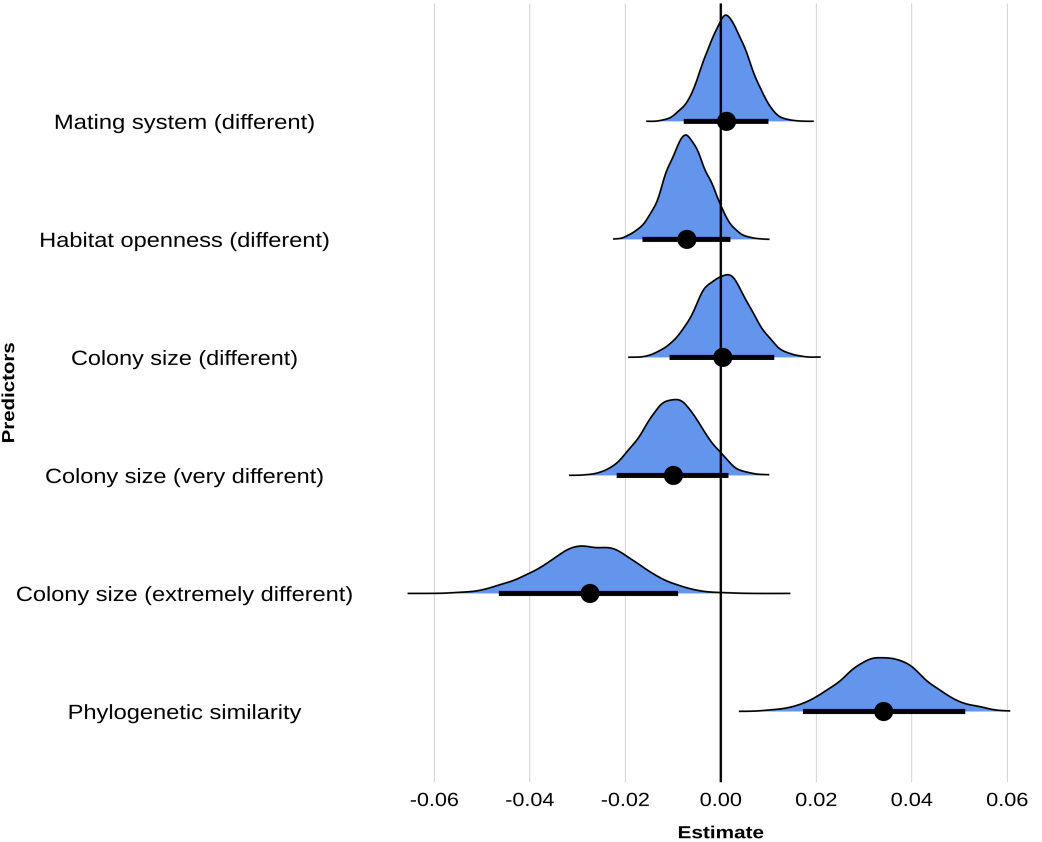
Figure 3 - Parameter estimates and posterior distributions from the bayesian phylogenetic multiple membership model performed on song composition (Mcomp). Estimates show a significant association between colony size and phylogenetic similarity and song composition similarity (response variable). The similarity in song composition between two species is lower when these species have marked differences in colony size. In other words, the greater the differences in colony size between two species, the more different their song compositions will be. The similarity in song composition is significantly lower when comparing two species with very different colony sizes compared to two species with similar colony sizes (reference level). Also, phylogenetic similarity is positively correlated to song composition similarity. The posterior distribution of the estimates of each of the tested predictors are shown with horizontal bars representing 95% of the distribution and the points indicating the means.
Discussion
Weaverbirds are widely known for their bustling colonies, where they can be observed parading around their intricately woven nests. While the acoustic properties and potential functions of their vocalizations have been studied in some species (Collias, 2000; Craig, 1976), the structure and diversity of their songs have not been investigated across species. The results of our comparative analysis showed that colony size and phylogenetic proximity predicted both song diversity and song composition. As expected, we found a positive effect of social complexity on these two song measurements. In contrast, no effect of mating system was observed. The significant association we initially found between mating system and song diversity in model Mdiv disappeared after completing the PPA, which models causal relationships. Lastly, phylogeny had a relatively high effect, explaining about one fifth of the song diversity variance. There was also a strong effect of phylogenetic proximity on song composition. Overall, these results show that, in weaverbirds, song evolution is constrained by phylogeny and affected by colony size. In contrast to what was commonly believed, sociality could exert a major influence on song evolution, even stronger than sexual selection. We discuss these results below.
Colony size as the main driver of song diversity and composition
In many mammal species sociality is associated with more complex vocalizations (Freeberg et al., 2012) and, in birds, cooperative breeding species have also been found to have a larger repertoire size for both calls and songs than non-cooperative ones (Leighton, 2017). Here, we found that, as predicted, weaverbird species living in larger colonies exhibit higher song diversity. As group size increases, the diversity of interactions between individuals probably also increases, thereby increasing the benefits of producing a larger variety of sounds for communication (Dale et al., 2001). It could also be that more diversity is needed to allow individual recognition in larger colonies (Briefer et al., 2008; see also Aubin & Jouventin, 1998). Thus, selection could operate on cognitive, motor, or morpho-anatomical traits that allow for the production of a wide diversity of sounds to facilitate communication. This selection could also be amplified by the need for significant cognitive abilities required for living in societies (Dunbar, 1998). However, if a correlation was found, it does not determine the direction of causality, and a greater diversity of signals might lead to larger colony sizes, a hypothesis that could be further explored through ancestral state reconstruction. It would also be interesting to examine, at the intra-species level, whether song diversity is greater in populations that live in larger colonies compared to populations living in smaller colonies.
Regarding song composition, our results are also in line with our prediction, supporting our hypothesis that species living in similar colony size have similar song syllables. It is likely that solitary species develop songs composed of syllables of a different nature compared to species living in large colonies because certain syllable categories could (i) be better perceived in noisy environments, as they might allow better discrimination of voice characteristics, (ii) lead to greater variability of syllables, which is useful for formulating various signals, or (iii) be easier to memorize, which could be advantageous as larger colonies tend to have more song diversity. In birds, two studies tested these mechanisms in social contexts beyond the recognition of partners, parents and descendants, or territorial neighbors and found that specific phonological and syntactical features facilitate individual recognition (Gentner & Hulse, 1998; Kondo et al., 2010).
As an extra note, we point out that even if the link between colony size and diversity suggests an effect of sociality, we cannot a priori exclude that larger colony size could be associated with stronger sexual selection because (i) mate choice is more intense as the encounter rate of mates is higher, the cost of rejecting mates is lower, more mates are available, and they can be more easily compared (Kokko & Rankin, 2006), and (ii) intrasexual competition is more intense and could drive a higher expression of signals (Diniz et al., 2018). Under this scenario, the effect of colony size that here is associated with sociality would be more closely linked to sexual selection. However, this conclusion would be at odds with the effect of the mating system that we discuss below.
No clear effect of mating system on song diversity and composition
We expected that stronger sexual selection acting on species would lead to greater song diversity. The intensity of sexual selection, here estimated by using the species’ mating system, was negatively correlated with song diversity when including colony size, habitat openness and mating system in the model (Mdiv). However, when considering causal effects between our studied variables and thus disentangling direct from indirect effects, the effect of mating system became non-significant, although polygamous species still present less song diversity than monogamous species. These results could be explained by the fact that weavers might experience no sexual selection on song diversity (at least in most species). Also, it is possible that other parameters of their songs are sexually selected, leading to a trade-off (Podos, 2022). In the context of intrasexual competition, rivals might focus on song parameters, other than diversity, that better predict the outcome of a fight or risk of injury. For instance, the repetition of the same syllable (Sierro et al., 2023) or the production of syllables or notes at a performance limit (DuBois et al., 2009) can indicate an individual's neuromotor capabilities or quality, which might be associated with its fighting capacity. Additionally, evaluating an individual's repertoire size can take time, making it a less relevant signal in territorial intrusion contexts (Rivera-Gutierrez et al., 2011). Another possibility is that our proxy for sexual selection is not capturing well the strength of sexual selection. The social mating system gives us an indication of the variance in male reproductive success. However, it would be important to incorporate extra-pair paternity data to see if the social mating system is close to the reproductive mating system in this family, but no data currently exist to evaluate this possibility. Finally, it is important to point out that mating system not only reflects the intensity of sexual selection but also a form of socialization. Indeed, monogamous species may exhibit stronger social bonds, where members of a pairbond must have a much finer understanding of each other’s needs and intentions (Dunbar & Shultz, 2010). For instance, a comparative analysis in primates showed that social bonds, measured by the time individuals spent grooming, were positively correlated with vocal repertoire size (McComb & Semple, 2005). Consequently, it is possible that this phenomenon explains why monogamous species have a greater diversity of songs than polygamous species.
Our study therefore concurs with recent interspecific studies that challenge the idea that increased sexual selection leads to increased complexity of song (Robinson & Creanza, 2019). At the interspecific level, few studies have explored the effect of mating system on acoustic diversity. The only two studies that examined this relationship using phylogenetic correction (with a sample size of 96 and 78 species) also did not find differences between polygamous and monogamous species (Snyder & Creanza, 2019; Hill et al., 2017a). Other proxies of sexual selection have been studied at the interspecific level. Price and Lanyon (2004) found a non-significant negative relationship between song versatility and sexual dimorphism in body size. Another study examined the relationship between the frequency of extra-pair paternity with three measures of song diversity (song and syllable repertoire size and versatility) and did not find a significant association (Garamszegi & Møller, 2004). Finally, a last study showed no correlation between sexual dichromatism and an ACP component loaded by three measures of diversity: number of syllables, number of syllable types, and syllable versatility (Ornelas et al., 2009). Taken together, our results and those of other studies suggest that there is no link between sexual selection intensity and diversity on a large scale.
We found that the syllable categories identified here are distributed equally among monogamous and polygamous species, which indicate that no types of syllables are more used when the intensity of sexual selection is stronger. This could be the case if there are no specific quality markers. In contrast, in a study performed on various families it was found that the song structure of polygamous species exhibits convergent acoustical displays (Loffredo & Borgia, 1986). In weaverbirds, we might find such patterns using more general categories of syllables, but we preferred to construct fine categories to maintain high discriminatory power and minimal arbitrariness.
Phylogeny constrains song evolution
Our estimation of the phylogenetic signal indicated that 18% of the variation of song diversity between species is explained by the phylogeny. This value can be compared to the Pagel’s λ that would indicate that song diversity partially evolved as a Brownian motion (Pearse et al., 2023; Housworth et al., 2004). This parameter indicates that phylogeny seems to play a role in explaining the variance of song diversity observed in weaverbirds, a result that aligns with other studies showing a significant effect of phylogeny on song complexity with Pagel’s λ equal to 0.86 and 0.84 for syllable repertoire size (Tietze et al., 2015; Snyder & Creanza, 2019). The presence of a phylogenetic signal in song parameters can lead to different interpretations. A recent study compared the phylogenetic signal of vocalizations produced by vocal learning in non-learning birds (Arato & Fitch, 2021) and found no differences between them. The authors concluded that the persistence of phylogenetic signal in learned vocalizations suggests that vocal learning is compatible with genetic determination (see also Price & Lanyon, 2002). This idea is supported by Blomberg et al. (2003) who assumed a direct link between low phylogenetic signal, evolutionary lability and heritability. However, at the interspecific level, the heritable component seems to include gradual genetic changes accumulated over the phylogeny but also any non-genetic response to an environment such as cultural evolution (Housworth et al., 2004). This suggests that, by interpreting the phylogenetic signal, we cannot assess if the similarity in song diversity between related species of weaverbirds came from a common cultural heritage or a common genetic heritage, while this objective could be achieved within a species (Wickler et al., 2006). In a recent study, Jablonszky et al. (2022) used a quantitative genetic method to estimate song versatility heritability in collared flycatcher and obtained a heritability of 0.09 (95%CI = [0.05, 0.13]) suggesting that birds mainly adjust their song to their current environment or condition, or that they learn their signals from other individuals than their parents.
Apart from telling apart the cultural and genetic effects, some authors use the phylogenetic signal to evaluate the strength of selection or the speed of evolution. For instance, by observing a weak signal on several acoustic parameters, Greig et al. (2013) suggested that these traits have probably been influenced by selection. However, other authors suggest that phylogenetic signal does not infer either selection or evolution rate (Revell et al., 2008; see Kamilar & Cooper (2013) for a review of the interpretation of the phylogenetic signal). Evolution rate of song parameters could however be interesting to quantify as it can indicate if the presence of cultural evolution as learned traits are subject to imprecise copying, which can quickly generate novel phenotypes (Mason et al. 2017 but see Medina‐García et al., 2015). These contradictions regarding the interpretation of the phylogenetic signal discourage us from inferring mechanisms of song diversity evolution in weavers. In this context, measuring the phylogenetic signal could only allow to control for phylogenetic relationship and show how much the trait we see is constrained by the phylogeny.
We also found a strong effect of phylogenetic proximity on song composition. Here again it can be related to either a genetic or non-genetic effects. This acoustic measure may be linked to syringeal and beak morphology or anatomy and neural control, themselves probably phylogenetically constrained. For instance, several studies support that beak morphology presents a phylogenetic signal and affects several acoustic parameters (Mejías et al., 2020; Huber & Podos, 2006; Podos, 2001 but see Porzio et al., 2019).
Conclusion
Our comparative analysis revealed an association between a proxy of social complexity, colony size, and song diversity and composition in the weaver family. Moreover, we found no clear association between a proxy for sexual selection, mating system, and song diversity and composition. While controversy surrounding the effect of sexual selection on song diversity persists at the intraspecific level, our results also suggest this effect is also not always present at the interspecific level. Our study emphasizes that the evolution of bird song is multifactorial and that it is important to consider social context when studying a trait that evolves in the context of sexual communication.
Acknowledgements
We would like to thank Flora Goujon, Camille Auneau and Mathieu Pelte for their help in labeling the songs, Pietro D’Amelio, Megan Thompson, Matthew Silk and Pierre de Villemereuil for their help on the statistical analyses, and Carrie Hickman and Kyle-Mark Middleton for having the patience to record red-headed weaver songs. Finally, we also thank PCI recommender Michael Greenfield and the two anonymous referees for their constructive comments.
Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Evolutionary Biology: https://doi.org/10.24072/pci.evolbiol.100836 (Greenfield, 2025).
Funding
The funding source of this study is French ANR through project 19-CE02-0014-01 Extended phenotype.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data, scripts, and supplementary information availability
Data, scripts and supplementary information are available online: https://doi.org/10.17605/OSF.IO/K7JR6 (Harscouet-Commecy, 2025).