
 CC-BY 4.0
CC-BY 4.0
Introduction
Fermented vegetables are traditionally consumed in central Europe and Asia, and have recently been the subject of renewed interest in Western countries, for many possible reasons that include greater consumer demand for more natural and sustainable foods and a growing proportion of vegetarian or vegan diets (Medina-Pradas et al., 2017; Thierry et al., 2023a). In Asian and Eastern countries, where fermented vegetables have formed part of traditional diets, a wide variety of vegetables are fermented and widely consumed (Gänzle, 2022; Thierry et al., 2023a). Cabbage is the main vegetable used worldwide, either in a mixture with other vegetables, as in Korean kimchi or Chinese paocai, or alone as sauerkraut in Eastern France and in Germany, where it is produced at an industrial scale (Tamang et al., 2020). In Western countries, sauerkraut, olives and cucumber are the main fermented vegetables that are commonly consumed. Other fermented vegetables are principally produced at the domestic and artisanal scales (Thierry et al., 2023a). In a recent study, we showed that French domestic and artisanal production concerns a wide variety of fermented vegetables. Indeed, within the framework of a citizen science project, the 75 samples collected from citizens included 23 types of vegetables, mainly cabbage (27%), followed by carrots (19%) and beets (12%), while 40% of them contained mixtures of vegetables (Thierry et al., 2023b). The potential health effects of plant-based fermented foods have only begun to be scientifically documented, even if they are being popularised by social media, generally without any scientific support (Thierry et al., 2023a). Some vitamin concentrations can increase, or be preserved, during fermentation but the effects depend on the microbial community and the conditions of production, among other factors, and contrasted results have been observed (Thierry et al., 2023a). All fermented vegetables are manufactured according to a relatively simple process, which consists in cutting and tightly packing raw vegetables with salt or brine, so that the vegetables are covered with brine or with the juices released from the vegetables (Buckenhueskes, 2015). Fermentation is usually spontaneous and due to an endogenous lactic acid bacteria (LAB) community (Buckenhueskes, 2015; Ashaolu & Reale, 2020). A wide variety of recipes are used in terms of the number, nature and mixture of vegetables, and the use of minor ingredients such as spices and condiments (Di Cagno et al., 2013; Ashaolu & Reale, 2020).
Several bacterial groups succeed each other over time during the spontaneous fermentation of vegetables, and some of them are alive at the time of consumption (Rezac et al., 2018). According to a meta-analysis of various fermented foods which covered 400 articles over 50 years, the average number of live microorganisms in fermented vegetable products such as sauerkraut, kimchi, pickles and olives ranges from 2 to 8 log colony-forming units (CFU)/g (Rezac et al., 2018). Environmental aerobic or facultatively anaerobic microorganisms grow first and are then gradually replaced by a succession of heterofermentative and then homofermentative LAB (Pederson & Albury, 1969; Buckenhueskes, 2015; Thierry et al., 2023a). For example, in a study on the spontaneous fermentation of carrot juice, bacteria from the Enterobacteriaceae family grew first to reach about 8 logCFU/g from the first hours of fermentation, and then decreased, disappearing totally after 10 days of fermentation (Wuyts et al., 2018). In parallel, during the first three days of fermentation, LAB actively grew, reaching around 9 logCFU/g and were responsible for a rapid drop in pH. The first LAB to grow are typically members of the Leuconostoc genus, followed by the Latilactobacillus and Lactiplantibacillus genera, with cell numbers reaching about 9 logCFU/g (Wuyts et al., 2018). Similar pictures have been observed with other vegetables, such as peppers (Li et al., 2024), sauerkraut (Müller et al., 2018) and cucumbers (Stoll et al., 2020). Most kinetic studies of fermented vegetables are carried out over relatively short periods of time and do not exceed one month, which is generally considered as the final stage of fermentation because the pH has stabilised (Wuyts et al., 2018; Müller et al., 2018; Wang et al., 2020). In a recent study carried out on 75 domestically-produced samples, 84% of them still contained live LAB, although the age of samples ranged from 2 weeks to 4 years with a median value of 6 months (Thierry et al., 2023b). LAB accounted for the majority of living microorganisms but also most of the 16S reads recovered by 16S rRNA gene metataxonomics while bile-tolerant Enterobacteriaceae were detected in only four samples (Thierry et al., 2023b). Alongside bacteria, yeasts and bacteriophages can also grow and survive in fermented vegetables (Tamang et al., 2016). Yeasts have been reported in various fermented vegetables (Liu et al., 2021; Wang et al., 2022a) and were found in half of 75 homemade fermented vegetable products analysed (Thierry et al., 2023b). Culture methods and culture-independent methods such as 16S metataxonomics are complementary as each method contributes specific information and potential biases (Parente et al., 2022) Culture methods enable quantification of the living share of the cultivable microorganisms present, while, by contrast, culture-independent methods provide access to all the microorganisms present in the sample, whether or not they are viable at the time of analysis. As for the 16S metataxonomics method, it is possible that combining several primers can overcome the specificity of the 16S primers used (Poirier et al., 2018a; Guo et al., 2022).
Some steps are essential to achieving successful fermentation, notably salting and packing. The main function of salting is to draw water and nutrients out from the vegetable tissue, thus supplying the microorganisms with the substrates they require for growth (Buckenhueskes, 2015). The NaCl concentration generally ranges from 1% to 3% of the final product (Buckenhueskes, 2015). Sliced vegetables are filled and pressed into vessels or glass jars, tight packing being crucial to eliminate air pockets and promote an anaerobic environment that will limit the growth of undesirable aerobic microbiota that might be responsible for spoilage (Buckenhueskes, 2015). The products are then allowed to ferment at ambient temperature for at least 3 to 4 weeks before being consumed or stored further at lower temperatures. These incubation conditions (temperature, NaCl concentration, oxygen availability, etc.) determine the start-up speed of the fermentation process and thus shape the microbial community (Thierry et al., 2023a). The rate of fermentation, and particularly the time course of pH decreases, is crucial to limiting the growth of undesirable microbiota (Buckenhueskes, 2015). Temperature and salt concentrations influence the dynamics of LAB species. The higher the temperature of fermentation, the more rapidly the pH will fall, and the earlier the dominance of (former) lactobacilli, which have a greater acid tolerance (Thierry et al., 2023a). In contrast, leuconostocs have been shown to be present at a higher abundance at temperatures <15°C (Wang et al., 2020). As for the salt concentration, early studies showed its impact on the rate of acidification and the growth dynamics within the main LAB species, e.g. the growth of Leuconostoc mesenteroides, which is less salt-tolerant than other, homofermentative, LAB species (Pederson & Albury, 1969).
The type of cutting markedly varies in a domestic production setting, as observed in a recent study (Thierry et al., 2023b), where vegetables were either cut thinly (grated, shredded), or more roughly (into slices, dices, or simply cut in two lengthwise, for example), or even left whole in the case of some small sized vegetables. For example, carrots can be thinly or more roughly grated, or cut into small dices or slices, or only roughly cut. However, to our knowledge, the role of this factor has only been rarely addressed. In an original study that explored the survival of inoculated Escherichia coli O157:H7 and Listeria monocytogenes during the fermentation of whole heads and shredded cabbage, both pathogens declined faster in shredded cabbage (Niksic et al., 2005). This was explained by the significantly higher total titratable acidity in shredded cabbage, compared to whole head cabbage. In summary, thin cutting is expected to facilitate the withdrawal of water and nutrients from vegetable tissue and thus to increase the buffering capacity of juice and accelerate the rate of (lactic acid) fermentation.
Our aim was therefore to investigate the effects of two factors, the cutting type and a slight reduction in the amount of salt added, on microbial and biochemical changes during the spontaneous fermentation of vegetables. We chose to study a root vegetable, carrots, and a leafy vegetable, cabbage, either thinly or roughly cut. We thus compared fermentation between grated carrot and sliced carrot, and between grated cabbage and whole cabbage leaves. We also studied two salt concentrations: a concentration of 1% (which is the minimum concentration of salt normally used), and, with a view to further reducing salt levels in line with health recommendations, a concentration of 0.8%. We performed the fermentations of carrot and cabbage under controlled conditions, and characterised the microbiological and biochemical changes in duplicate (two independent jars) over one month by combining culturomics, 16S rRNA gene and gyrB metataxonomics analysis for bacterial communities, and targeted metabolomics.
Materials and methods
Ingredients and experimental design
Two vegetables, carrot and white cabbage, were collectively chosen by partners in the FLEGME citizen science project as being among the most frequently used in the manufacture of fermented vegetables, so as to include a root and a leafy vegetable. Unwashed organic carrots were supplied by the Ty Coz farm in Saint-Pol-de-Léon, France. Unwashed organic white cabbages (Brassica oleracea L.), were supplied by the Coopérative des Producteurs Légumiers, Doué en Anjou, France. Dry (<0.1% humidity) grey coarse sea salt and tap water were used to prepare the brine. 500 g-jars with glass lids and rubber seals were used for storage (Korken, IKEA).
A 23 experimental design was set up with (i) vegetable type (carrot/cabbage), (ii) cutting type (thin/rough) and (iii) salt concentration (0.8%/1%) as the factors. The two vegetables, cabbage and carrot, were thinly or roughly cut, and then firmly pressed down in 500 mL-jars which were filled up with brine to reach final NaCl concentrations of 0.8% or 1.0%, expressed as gram of raw salt per 100 g of the vegetable and brine mixture (Figure 1). More precisely, the carrots were either grated or cut into slices, and the cabbage leaves were shredded or cut into pieces measuring about 6-8 cm each way.
After removing the external leaves, the cabbages were either shredded using a professional Dito Sama TRS vegetable slicer equipped with a 2 mm disk, or the leaves were cut manually into ~6 cm x ~8 cm pieces. Then, 205 g shredded cabbage and 282 g brine, or 232 g cabbage leaf and 246 g brine, were weighed into each jar. As for the carrots, they were washed, hand-peeled, grated in 3 mm pieces or cut into 5 mm slices at the CTCPA pilot facility (Agri-food Technical Centre and Oniris, Nantes, France). 285 g grated carrot and 215 g brine, or 285 g sliced carrot and 230 g brine, were then weighed into each jar. To ensure the final expected salt concentrations of 0.8% and 1%, preliminary tests were performed to determine the maximum amount of vegetables that could be packed in a jar, which ranged from 205 g to 285 g depending on the vegetable and cutting type (Supplementary Table S1), and, consequently, the quantity of liquid (brine) that could be added (215 to 282 g), in order to calculate the salt concentration of the brine in each case. The details of brine concentrations are given in Supplementary Table S1.
Twenty jars of each type (vegetable, cutting type, salt concentration) were prepared, leading to a total of 160 jars. These jars were transported (1 h of transport between the manufacturing site and laboratory) at ambient temperature (approx. 20°C) just after manufacture in order to be incubated at the STLO laboratory at 19°C for 7 months. Two independent jars were characterised at each time point. One sample of raw vegetable was transported in a cooler (approx. 6°C on arrival) to be analysed before fermentation (T0).
The samples were characterised for viable microorganisms, metataxonomic profiles and pH measurements at four sampling times. The first sampling time was at the very beginning of fermentation; two samples (named T1 and T2) then covered the initial acidification period and the last (T3) was collected after four weeks of incubation. Due to differences in the acidification rates, the first two sampling times were 40 h (T1) and 64 h (T2) for carrot and 40 h (T1) and 86 h (T2) for cabbage. A late sampling (T4) was performed after seven months of incubation, for microbial enumeration and pH measurement only (Figure 1). In addition, some intermediate samples were collected between T2 and T3 for volatile analysis and isolate identification (T2a, T2b, and T2c: 10, 15 and 21 days for carrot samples and T2b and T2c: 14 and 22 days for cabbage).
The sample names were coded as follows: 301 and 311 for shredded cabbage at 0.8% and 1.0% salt, respectively, 321 and 331 for cabbage leaf at 0.8% and 1.0% salt, respectively, 401 and 411 for grated carrot at 0.8% and 1.0% salt, respectively, and 421 and 431 for sliced carrot at 0.8% and 1.0% salt, respectively (Figure 1). For example, sample 331-a-T3 was the replicate ‘a’ of a cabbage leaf sample prepared with 1.0% salt, sampled after 4 weeks of fermentation.
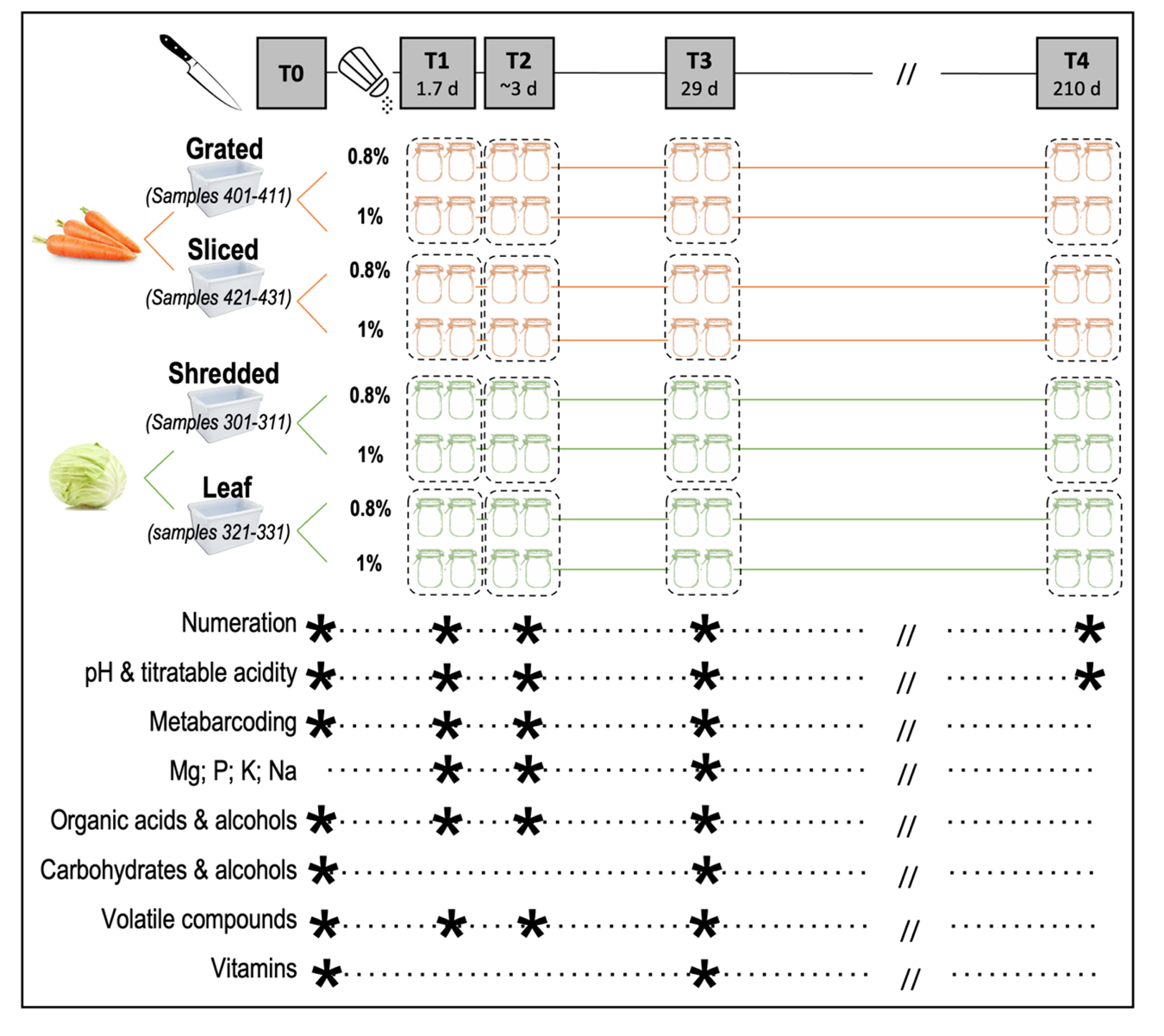
Figure 1 - Experimental design used to prepare fermented vegetables and sampling. The three factors tested were vegetable type (carrot and cabbage), cutting type (thin or rough) and salt concentration (0.8% and 1%, expressed as g raw salt per g of preparation (vegetable and brine). Sampling was performed in duplicate (two independent jars) at each sampling point, represented by a star, except for volatile analysis and vitamins, performed in triplicate (three independent jars), at five time points: T0, initial time, T1, 1.7 day, T2, 2.7 (carrot) or 3.6 day (cabbage), T3 (4 weeks), and T4 (7 months). Some analyses were also performed at 9 days (stage T2a, carrot only), at 2 weeks (stage T2b), and at 21 days (stage T2c, cabbage only).
Culturomic conditions
Samples of 10 g fermented vegetables (5 g juice plus 5 g drained vegetables) were suspended in 90 mL of a Tryptone Salt diluent (TS, sodium chloride 8.5 g/L, tryptone 1 g/L) heated at 48°C and homogenised in a filter bag (BagPage+, Interscience) in which the vegetable debris was separated from the filtrate. Microbial analyses were performed on 14 different nutritive and selective media and incubated under aerobic (air atmosphere) or anaerobic conditions (Anaerocult® A, Merck, Darmstadt, Germany) at 37°C, 30°C or 25°C depending on the medium, as previously detailed (Thierry et al. 2023b). In brief, seven media targeted the following microbial groups: LAB, total aerotolerant bacteria, halotolerant bacteria, aerotolerant Gram-negative bacteria, yeasts and filamentous fungi, bile-tolerant Enterobacteriaceae, and enterococci (Supplementary Table S2). In addition, three media targeted spore-forming bacteria (Supplementary Table S2). Bacillus cereus-typical colonies on BCA were further examined, by observing their aspect on the Compass Bacillus cereus agar medium (Biokar), incubated at 30°C for 24 h and 48 h, microscopic observations and panC gene sequencing.
In addition, four pathogens, namely Escherichia coli, coagulase-positive staphylococci (Staphylococcus aureus), Salmonella, and Listeria monocytogenes, were searched for by a subcontracted laboratory (LABOCEA, Fougères, France) according to ISO 16649-2, ISO 6888-2, BRD 07/11-12/05 and AES 10/03-09/00 standards, respectively.
Microbial isolation and identification
To collect LAB strains, one to three isolates were picked up on the several culture media used for agar plates containing 20 to 100 colonies, according to visual aspect of the colonies (size, colour, morphology), in order to favour the diversity of the isolates collected. Yeast isolates were collected using the same methodology. The isolates were collected from T0 to T4 with an intermediate collection stage at 15 days, corresponding to the stabilisation of pH. Bacteria and yeast clones were identified using the 16S rRNA gene and the D1/D2 domain of 26S rRNA gene sequencing, respectively. Bacteria and yeast were identified by the 16S rRNA gene and the D1/D2 domain of 26S rRNA gene sequencing, respectively.
16S rRNA gene and gyrB metataxonomic analysis
DNA was extracted from the samples using the Nucleospin Tissue kit (Macherey-Nagel, Düren, Germany) as previously described (Thierry, 2024). DNA sequences were amplified in the 16S rRNA gene V5-V7 region for bacteria using 799F/1193R primers (Forward- AACMGGATTAGATACCCKG, Reverse-ACGTCATCCCCACCTTCC) and under the PCR conditions previously described (Beckers et al., 2016). In parallel, the degenerate primers F64 (5’-MGNCCNGSNATGTAYATHGG-3’) and R353 (5’-CNCCRTGNARDCCDCCNGA-3’) were used to amplify a ~280-bp region of gyrB (Poirier et al., 2018b). The 16S rRNA and gyrB amplicons were sequenced at the Génome Quebec sequencing platform (Montreal, Quebec) using Illumina MiSeq PE250 technology, which generated 2 x 250 bp reads and a total of 2.45 Gb of data for amplicons.
Bioinformatic analyses
The raw sequences of 16S rRNA gene sequencing were processed as previously described (Thierry et al., 2023b). The raw sequences of gyrB gene sequencing were also processed using the DADA2 package v 1.20.0 (Callahan et al., 2016), following the authors guidelines: we successively applied the filterAndTrim, learnErrors, dada, mergePairs, makeSequenceTable, removeBimeraDenovo and assignTaxonomy functions. The gyrB database from (Poirier et al., 2018b) was used to determine taxonomic affiliation. The amplicon sequence variants (ASV) count table, the ASV taxonomy table and the sample metadata were combined into one phyloseq object for each target gene. The phyloseq (v1.44) R package (McMurdie & Holmes, 2013) was used to visualise barplots. Data were transformed into relative abundances before computing beta diversity (Bray-Curtis dissimilarity). The ComplexHeatmap (v2.16.0) R package (Gu et al., 2016) was used to visualise the relative abundance of the different genera on a heatmap with a complete clustering based on Bray-Curtis dissimilarity computed after depth normalisation. Principal coordinate analyses (PCoAs) based on Bray-Curtis dissimilarity, calculated from the relative abundances of different genera, were conducted to evaluate the beta diversity of the samples using gyrB and 16S markers.
Biochemical analyses
The pH of juice samples was measured with a pH-metre (Hanna Instruments HI 2020-02).
Total titratable acidity (TTA) was determined on centrifuged (18,000 g for 10 min at 20°C) juice samples, by titrating approximately 10 mL juice with 0.1 M NaOH to pH 8.3. It was estimated as follows: total acidity (%) = VNaOH × 0.1 × m / S /10, with VNaOH, volume of 0.1 M NaOH (mL); 0.1, factor corresponding to NaOH normality; m = 90, molar mass of lactic acid, S, mass of sample used (g). TTA was expressed as a percentage (w/w) of lactic acid.
Sample preparation
Before metabolite analysis (except for volatiles and vitamins), aliquots of juices were first centrifuged at 8000 g for 10 min at 4°C to eliminate plant debris and the supernatant was deproteinized by ultrafiltration on Vivaspin 2 centrifugal concentrator columns (polyethersulfone, 10 kDa cut-off, Sartorius) at 8000 g for 15 to 30 min at 4°C. Two chromatographic systems: High-Performance Liquid Chromatography (HPLC) coupled to UV and refractometry detection, and high-performance anion-exchange chromatography coupled to pulsed amperometric detection (HPAEC-PAD), were combined to analyse a range of organic acids, carbohydrates and alcohols.
Before mineral analysis, the juice samples were centrifuged at 18,000 g for 10 min at 4°C, and the supernatant 40-fold (for Mg and P analysis) to 1000-fold diluted (for Na and K analysis) in a 2% v/v HNO3 (Thermo fisher scientific, Waltham, MA, USA).
For volatile metabolites, triplicate samples taken from three independent jars were directly analysed from juice using headspace (HS) gas chromatography-mass spectrometry (GC-MS). Juice aliquots (2.5 mL) were placed in Perkin-Elmer 22 mL vials (B0104236, 20 mm) and hermetically sealed, and the vials were stored at -80°C until analysis.
Vitamin analysis was performed in drained vegetable samples stored at -20°C before analysis.
Acids and alcohol analysis by high-performance liquid chromatography
Supernatants were 2- to 4-fold diluted in 0.005 mol-L-1 H2SO4 and stored at -20°C until analysis. Lactic, acetic, citric, succinic, oxalic and pyruvic acids, ethanol and butanediol were quantified by High-Performance Liquid Chromatography (HPLC, Ultimate 3000, Thermo Fisher Scientific 91941 Courtaboeuf, France), using a Rezek ROA organic acid H + column (300*7.8 mm, Phenomenex, California), with H2SO4 0.005 M as the mobile phase at a flow rate of 0.4 mL/min at 60°C. Two detectors were used: a UV detector (DIONEX-UVD 1704) operated at 210 nm and a refractometer (RI 2031 Plus Jasco).
The data were processed using Chromeleon™ software. Quantification was performed using multi-standard external calibration. Standards of ethanol, butanediol, oxalic, lactic, citric, propionic, butyric, succinic and pyruvic acids were obtained from Merck, St. Quentin Fallavier, France, and acetic acid from PanReac, Lyon, France. Mannitol, fructose and glucose can also be analysed using this method but in the present study were not quantified because the sucrose of vegetables hydrolyses in glucose and fructose during analysis, and fructose is co-eluted with mannitol.
Carbohydrate analysis by high-performance anion-exchange chromatography
The supernatants were diluted 400-fold in milli-Q® water (Merck, Darmstadt, Germany) and kept frozen at -20°C until analysis. Carbohydrates (sucrose, glucose, fructose, galactose, raffinose, xylose, arabinose, mannose, and mannitol) were quantified by high-performance anion-exchange chromatography (HPAEC) and pulsed amperometric detection (PAD) on an ICS-5000+ Dionex system (Thermo Electron SAS, Courtaboeuf, France), as previously described (Canon et al., 2020). The system was equipped with a Dionex CarboPac PA210-Fast-4 µm column preceded by a CarboPac PA210-4 µm guard column (2*30 mm). Metabolites were eluted with KOH as the eluent, at a flow rate of 0.2 ml/min with the following gradient: 0 to 32 min 13 mM, 32 min to 55 min 100 mM then return to 13 mM from 55 to 65 min. Data were acquired and processed using Chromeleon7™ software (Thermo Scientific). Metabolites were quantified using multi-standard external calibration (prepared at 0.1 mg/L to 40 mg/L (Merck, St. Quentin Fallavier, France).
Mineral analysis using inductively coupled plasma-optical emission spectrometer (ICP-OES)
Minerals were quantified in the initial brine and in juices during incubation using an inductively coupled plasma-optical emission spectrometer (ICP-OES) (iCAP 7200, Thermo Fisher Scientific, Courtaboeuf, France), as previously described (Martin et al., 2022). Sodium, potassium, magnesium, phosphorus and selenium were quantified using standard external calibration. Na, Mg, K, and P standards were prepared from 100 ppm standard solutions (Reagecon, Shannon, Ireland) at 0.5 to 10 ppm in a 2% v/v HNO3 solution, and Se at 0.01 to 1 ppm in a 2% v/v HNO3 solution. NaCl concentrations were calculated from Na concentrations.
Volatile analysis by headspace - gas chromatography – mass spectrometry
Volatile compounds were extracted using a Turbomatrix HS-40 trap automatic headspace sampler and analysed using a Clarus 680 gas chromatograph coupled to a Clarus 600T quadrupole mass spectrometer, operated within a mass range of m/z 29 to m/z 206 and an ionisation impact of 70 eV (Perkin Elmer, Courtaboeuf, France) as previously detailed (Pogačić, 2015). Volatiles were eluted on an Elite WAX ETR column (30 m by 0.25 mm by 0.25 mm; Perkin Elmer, Waltham, MA), with helium as the mobile phase, under the following conditions: initial temperature 35°C maintained for 10 min, then increased to 5°C/min up to 230°C. Volatiles were identified by comparing their mass spectra and retention index with data from the NIST 2008 Mass Spectral Library data (Scientific Instrument Services, Ringoes, NJ, USA), from the literature and from standard injections, when available. Volatiles were semi-quantified from the abundance of one specific mass fragment (m/z), in arbitrary units. Mass spectrometry (MS) data were processed using XCMS on R software (R Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria). The full width at half maximum was set at 5, the maximum number of peaks per ion at 1000, the interval of m/z value for peak picking at 0.4, the signal-to-noise ratio threshold at 6, the group bandwidth at 3 and the minimum at 0.4. The other parameters were those by default.
Vitamins
Vitamins C, K1 and B9, and for carrot, beta-carotene, were quantified in vegetable samples (without juice) sampled at T0 and T3, while vitamins K2 and B12, which are not present in raw vegetables but potentially produced by bacteria, were analysed in T3 samples only. Vitamin C was analysed at the Vegenov laboratory (Saint-Pol-de-Léon, France), vitamins B9 et K1 by a subcontractor laboratory (Labexia, Quimper, France), and beta-carotene and vitamins B12 and K2 were determined by another subcontractor laboratory (Agrobio, Bruz, France), using HPLC and LC-fluo internal methods.
Vitamin C was determined in accordance with the NF V03-135 standard. Briefly, vitamin C was extracted from samples using a 20 g/l metaphosphoric acid solution. L(+)-dehydroascorbic acid was reduced in L(+)-ascorbic acid using a 40 g/l L-cysteine solution. L(+)-ascorbic acid was quantified by HPLC (Agilent, Les Ulis, France) with a photo diode array at 265 nm as the detector.
Vitamin B9 was extracted from the samples, and the diluted extracts and test broth medium were placed in the wells of a Vitafast B9 microplate kit, in the presence of a Lacticaseibacillus rhamnosus strain, and incubated in the dark at 37°C for 44 to 48 h. The intensity of the metabolism of L. rhamnosus due to the vitamin B9 supplied by the extract was measured by turbidity using a microplate reader at 620 nm and compared to a standard curve.
Vitamin K1 was determined according to the NF EN 14148 standard. Fat was first eliminated by an enzymatic treatment, and vitamin K1 was measured by HPLC with fluorescent detection, after post-column reduction.
Statistical analyses
Three-way ANOVAs were performed for each vegetable to determine whether the microbial and biochemical variables differed according to the fermentation stage, cutting type, NaCl concentration and the 2-way interactions, using the R function aov. Means were then compared using the sidak posthoc test from the R package emmeans (Lenth, 2024).
In Figures 2 and 4, means and 95% confidence intervals were calculated using the excel functions AVERAGE and CONFIDENCE.NORM, respectively. In both these figures, we gathered the four values corresponding to the duplicate jars at the two salt concentrations, because either no (cabbage) or limited (carrot) effects of salt concentration were observed.
Principal component analyses (PCA) were performed using the PCA function of the FactoMineR R package (Lê et al., 2008). PCA was performed to illustrate the global biochemical and microbiological composition of the vegetables during fermentation and the relationships between the different variables. The 15 active variables were the pH and TTA values, the concentrations of the six main metabolites and the data from the enumeration of seven viable microbial groups in 56 samples (cabbage and carrot analysed at four fermentation stages). A further PCA was also performed on the volatile profile for each vegetable.
A multi-block Partial Least Squares-Discriminant Analysis (PLS-DA) was performed to determine if samples exhibited different signatures at T3 regarding metataxonomics (16S and gyrB), metabolites (lactic acid, acetic acid, mannitol, ethanol, butanediol), volatiles and enumeration results, given their cutting type or salt concentration. The relative abundance values of a genus were retained only for the marker in which they were the highest. The block.plsda function of the R package mixOmics (Rohart et al., 2017) was used with two components (ncomp = 2). This method was chosen for its ability to model heterogeneous, multi-block data, allowing for the identification of the best variables that discriminated the samples. Outputs were visualised with the plotIndiv and plotVar R functions, and the discriminating power of the model was interpreted thanks to the auroc R functions, giving the AUC and Wilcoxon test p-values for each class comparison performed.
Results
Establishment of the microbial community: Enterobacteriaceae preceded lactic acid bacteria
The time-course of growth of the two main bacterial groups, i.e. bile-tolerant Enterobacteriaceae and LAB, the pH decrease and total titratable acidity (TTA), is depicted in Figure 2 during the first month of fermentation. The fermentation stage significantly (p-value <0.01) impacted all these variables (Supplementary Table S3). The first bacterial group that grew was bile-tolerant Enterobacteriaceae, enumerated on VRBG and referred to as ‘enterobacteria’ below. The initial numbers of enterobacteria were 4.3 and 6.0 logCFU/g, in carrot and cabbage, respectively, after which they rapidly increased to reach about 7 and 8 logCFU/g in the cabbage and carrot samples, respectively (Figure 2). LAB were present at low numbers in raw carrot (3.16 +/- 0.06 logCFU/g), whereas they were below the detection level in cabbage. LAB grew after enterobacteria and reached maximal numbers of about 8 and 9 logCFU in cabbage and carrot, respectively. Identification of the 58 isolates collected from MRS agar medium, used to target LAB, effectively showed only LAB isolates. Simultaneously with LAB growth, the pH decreased and total titratable activity (TTA) increased. The pH in raw cabbage was 6.3, falling to 3.9 in about 2 weeks. In carrot, the pH fell from 5.8 to 3.8 in about 40 h in all samples. Afterwards, the pH only very slightly decreased, to reach 3.44 ± 0.18 and 3.39 ± 0.30 in carrot samples after 1 and 7 months of fermentation, respectively, and 3.94 ± 0.21 and 3.55 ± 0.13 in all cabbage samples after 1 and 7 months, respectively (results not shown at 7 months). TTA increased to 0.6% in cabbage and 1.1% in carrot.
Marked differences in microbial counts, pH and TTA values were observed between the duplicate jars sampled at each time point, but the results of ANOVA nevertheless highlighted some significant trends concerning the effect of the factors tested, i.e. cutting and, to a lesser extent, salt (Supplementary Table S3).
In cabbage, a global effect of cutting, but not of the salt concentration, was observed. In thinly-cut cabbage, LAB grew more rapidly to reach 3 logCFU more at T2 when compared to roughly-cut cabbage (p-value of stage*cutting factor: 0.013, and LAB mean counts of 7.5 and 4.6 logCFU/g in shredded and leaf cabbage, respectively). LAB counts reached 8 logCFU/g after ~5 and ~13 days in shredded and leaf cabbage, respectively, then decreased more rapidly in the former. Consecutively, the pH fell more rapidly in shredded cabbage (p-value <0.01), with a difference of 0.47 at stage T2. Accordingly, enterobacteria counts tended (p-value=0.07) to decrease faster in shredded cabbage, in which no enterobacteria were detected after 14 days of fermentation, while they were still detected in the half-leaf cabbage samples after fermentation for one month, despite the pH falling to between 3.2 and 4.4 (Figure 2). At that time, three out of the four isolates from the VRBG medium were identified as Hafnia alvei. Viable enterobacteria were, however, no longer detectable in 7-month old cabbage, in which the pH was 3.1 to 3.7 (data not shown).
A more complex picture was observed regarding carrots. Enterobacteria counts were significantly affected by the cutting type (p-value <0.001) but not by the salt concentration. They were lower in grated carrot than in sliced carrot at all stages, with a difference of up to 2.8 logCFU less at T2. Regarding LAB, they tended to grow more rapidly in grated carrot, with 0.5 logCFU more at the very start of fermentation compared to sliced carrot, and then to survive better in sliced carrot with 2.0 logCFU more at T3 (p-value of stage*cutting factor: 0.10). They also significantly (p-value salt factor = 0.04) survived better in 1%-salted samples, with about 2 logCFU more from T2. Consistently with LAB numbers, both the pH and TTA values were significantly affected by the cutting type and salt concentration, and differently depending on the fermentation stage (p-values of stage*cutting factor <0.001 and stage*salt factor <0.001). Hence, at T2, grated carrot had a lower pH and a higher TTA than sliced carrot (pH 3.69 vs pH 3.86 and TTA 0.76 vs 0.32, respectively) while the reverse was seen at T3. In addition, the 1%-salted carrot samples at T3 had a lower pH and a higher TTA than the 0.8% salted samples, regardless of cutting type (pH 3.30 vs pH 3.58 and TTA 1.29 vs 0.83, respectively).
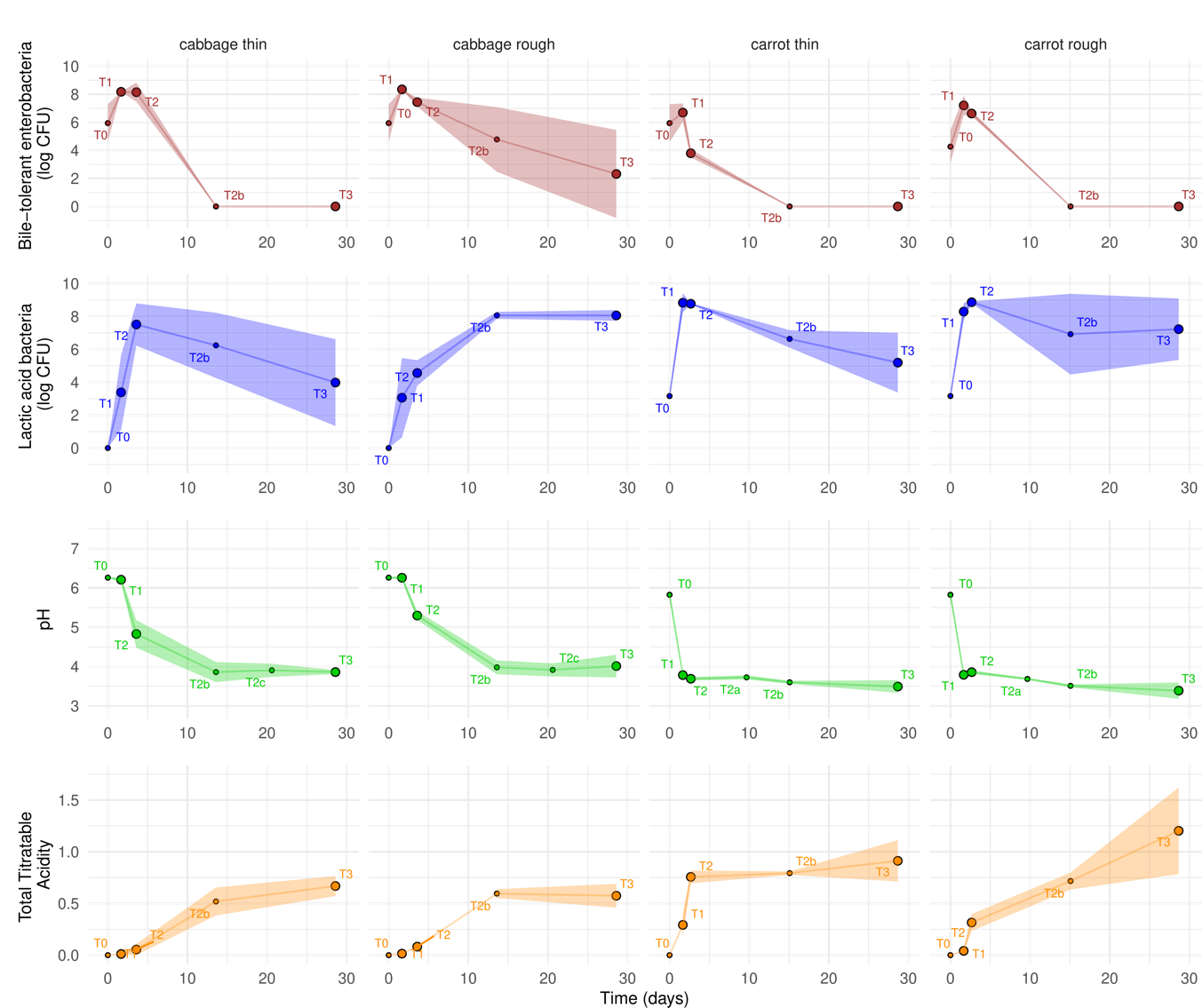
Figure 2 - Time-course of fermentation in cabbage and carrot for each cutting type, illustrated by the change in viable counts of bile-tolerant Enterobacteriaceae, enumerated on VRBG and lactic acid bacteria enumerated on MRS, expressed as logCFU/g, pH and total titratable acidity (TTA). Values are means of the results observed in two to four independent jars, and the size of the symbols is proportional to the number of replicates (n=2 at T0, T2a, T2b, T2c and n=4 for the other points, where the four values corresponded to the duplicate jars at the two salt concentrations, gathered because either no (cabbage) or limited (carrot) effects of salt concentration were observed. The coloured areas show the 95% confidence intervals.
Enterococci, selectively enumerated on KF medium, were detected at low counts (<4 logCFU/g, i.e. less than 0.04% of total LAB) at the beginning of fermentation in both vegetables. They disappeared after two weeks of fermentation, except in the leaf cabbage samples, which contained from 1.8 to 8.3 logCFU enterococci/g at one month.
Yeasts were present on the raw vegetables at a rate of about 2 logCFU/g. They did not grow in cabbage, from which they disappeared within 4 days (stage T2) in roughly-cut cabbage, faster than in thinly-cut cabbage where they were still 2.45 logCFU at T2 (p-values of stage factor: 0.0004, cutting factor: 0.0012 and stage*cutting factor: 0.015). By contrast, yeast counts did not significantly vary over time and as a function of the cutting and salt factors. However, yeasts grew in half of the carrot jars, with marked variations between the duplicates; for example, 0 and 7.9 logCFU/g in the 1% salt carrot duplicates at T3.
As for undesirable bacteria, none of the four pathogenic bacteria targeted were detected. Some spore-forming bacteria were present at low counts in carrot samples (median value of 75 CFU/g enumerated on a rich medium, BHI-YE) and were absent from cabbage samples except for one, a leaf cabbage sample (331-a-T3). This sample was also the only one that contained clostridia (4 logCFU/g, enumerated on the selective TSN medium), and also 2.44 logCFU/g Bacillus on BCA medium.
A total of 191 bacterial strains were thus isolated from carrot and cabbage between T0 and T3, and identified to the species level. Leuconostoc was the main LAB genus identified and represented 52% of the isolates collected during the first 15 days of fermentation, followed by Enterococcus (22%) and Lactiplantibacillus (10%) genera (Supplementary Table S4). Concerning non-LAB isolates, Bacillus dominated, followed by Hafnia, Pantoea, Rahnella, and Pseudomonas isolates. The 13 yeast isolates identified were members of Kazachstania, Rhodotorula, Saccharomyces, Candida, Pichia, and Debaryomyces (Supplementary Table S4).
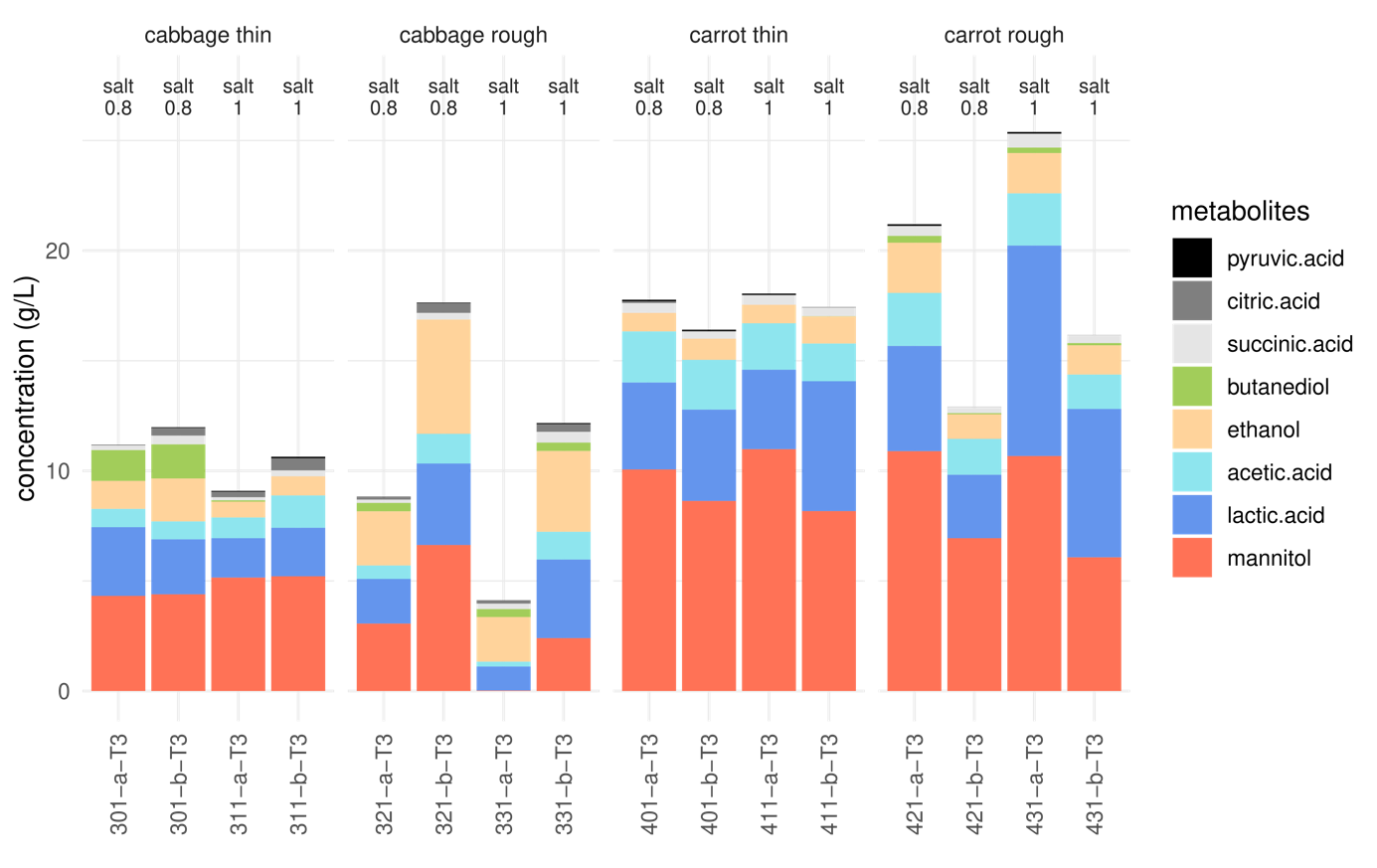
Figure 3 - Levels of metabolites in fermented cabbage and carrot juice, expressed in g/L, after one month of fermentation (stage T3), for each cutting type, and two salt concentrations, in two replicate jars coded a and b. Sample codes: 321, 301, 421 and 401 show samples at 0.8% and 331, 311, 431 and 411 at 1.0% salt.
Mannitol and lactic acid were the principal metabolites produced
Carbohydrates, organic acids and alcohols were analysed in the sample juices. In terms of carbohydrates, saccharose, glucose and fructose were the principal compounds detected at the start of fermentation (T0). After one month of fermentation (stage T3), the main vegetable carbohydrate detected in both vegetables was glucose, with marked variations in concentrations (0 to 4.8 g/kg juice), followed by saccharose in carrot (0.5-1 g/kg juice) and fructose (0 to 0.4 and 0.1 to 1.6 g/kg juice in carrot and cabbage, respectively). Small quantities of galactose (~250 mg/L) were also detected at T3 in both vegetables, and traces (<30 mg/L) of xylose, mannose, and arabinose, and, of raffinose in cabbage only.

Figure 4 - Time-course (days) of fermentation in cabbage and carrot for each cutting type, illustrated by changes to the levels of lactic acid, acetic acid and ethanol, in g/kg juice. Values are means of the results observed for two to four independent jars, and the size of symbols is proportional to the number of replicates (n=2 at T0 and T2a, T2b, T2c and n=4 for the other points) where the four values corresponded to the duplicate jars at the two salt concentrations, grouped because no significant effect of salt concentration was observed on cabbage. The coloured areas show the 95% confidence intervals, with either no or limited (carrot) effects of salt concentration being observed.
The main metabolites were mannitol and lactic acid, followed by acetic acid, which together accounted for 77% to 92% of total metabolites at T3, except in the roughly-cut cabbage samples, which also contained high levels of ethanol (Figure 3). The other minor compounds detected were 2,3-butanediol (up to 0.31 g/kg in carrot and 1.5 g/kg in cabbage, respectively, at T3), succinic acid (~0.35 g/kg juice), citric acid (~0.30 g/kg cabbage juice), oxalic acid (~0.07 g/kg carrot juice), and pyruvic acid (~ 0.04 mg/kg juice). It is worth noting that there were large differences, mostly quantitative, were observed at T3 between the duplicate jars (Figure 3).
The concentrations of all metabolites except for butanediol rose significantly over time (p-value <0.01, Supplementary Table S3). Their concentrations stabilised in juice after ~3 days in grated carrot, ~2 weeks in sliced carrot and shredded cabbage, and ~30 days in cabbage leaf, as illustrated in Figure 4 for lactic acid, acetic acid and ethanol.
In cabbage, the time-course of lactic acid and acetic acid production did not vary significantly as a function of the cutting and salt factors (Figure 4). Their mean concentrations at T3 were 2.5 and 0.9 g/kg, respectively (Figure 3). By contrast, ethanol production, was significantly affected (p-value <0.001) by the cutting type, with concentrations of 3.3 and 1.2 g/kg in leaf and shredded cabbage, respectively (Figures 3 and 4).
As previously noted, the picture was more complex in carrots, with significant effects of cutting, of the interaction cutting*stage, and/or interaction cutting*salt, depending on the variable (Supplementary Table S3). Globally, lactic and acetic acid were produced more rapidly at the start of fermentation in grated carrot (e.g. 0.15 and 2.40 g lactic acid /kg and 0.11 and 1.33 g lactic acid /kg at T1 in sliced and grated carrot juice, respectively). By contrast, ethanol was produced at significantly higher levels (p-value <0.001) in grated carrot, with mean values of 0.97 and 1.64 g/kg at T3 in grated and sliced carrot juice, respectively (Figures 3 and 4).
PCA highlighted the global changes to microbial and biochemical composition over time and the impact of cutting
PCA was performed to globally illustrate the effects of the cutting and salt factors on microbial and biochemical changes during fermentation. A total of 15 variables were used: the viable counts of microorganisms (n=7), the amounts of the main metabolites (n=6), and the pH and TTA values, for all cabbage and carrot samples analysed over time (Figure 5A and 5B). The first axis, which explained 42.2% of total variability, clearly separated samples on the basis of the time-course of fermentation (Figure 5C). The pH value, enterobacteria counts and Gram-negative aerotolerant bacteria (tot_aero_Gneg, enumerated on BHI-YEnp) were negatively associated with PC1 and associated with the start of fermentation (Figure 5B), as shown above (Figure 2). Nearly all of the isolates (28 out of 30) collected from BHI-YEnp medium at up to 15 days fermentation were members of the Enterobacterales order (Enterobacteriaceae, Hafniaceae, Erwiniaceae, Yersiniaceae family) (Supplementary Table S4). In contrast, the levels of most metabolites and TTA were positively associated with PC1 and characterised 2- to 4 week-fermented samples (Figure 5A and 5B), as shown above (Figures 2 and 4). The second axis, which explained 23.2% of total variability, was associated with intermediate fermentation stages and separated samples on the basis of high viable counts in three groups of Gram-positive bacteria: i) LAB, ii) halotolerant bacteria enumerated on TSA-NaCl medium from which 11 out of the 14 isolates collected at T3 were identified as LAB, and iii) total aerobic bacteria (total_aero_bacteria) enumerated on the BHI-YEn medium from which Pseudomonas sp. and Janthinobacterium sp. isolates were identified at T0, while only LAB were isolated from T2 (Supplementary Table S4). Each vegetable displayed a specific course of changes from T1 to T3, depending on the cutting type, as shown by the lines linking the successive stages on the PCA individual graph (Figure 5A). Thinly-cut cabbage, shown in green, had a similar composition at T1 compared to roughly-cut cabbage, shown in dark green, but displayed more rapid lactic fermentation, i.e. an increase in LAB counts, and decreases in pH and enterobacteria counts, as described above (Figures 2 and 4). Concerning carrot, differences between thinly- and roughly-cut carrot (shown in orange and red, respectively) were perceived as soon as stage T1, with the former exhibiting a higher rate of fermentation, as stated above (Figures 2 and 4). Similar profiles were however observed for all carrot samples from two weeks of fermentation (T2b and T3). These global differences were also clearly shown according to the vegetable and cutting type, but not to the salt concentration (Figure 5D).
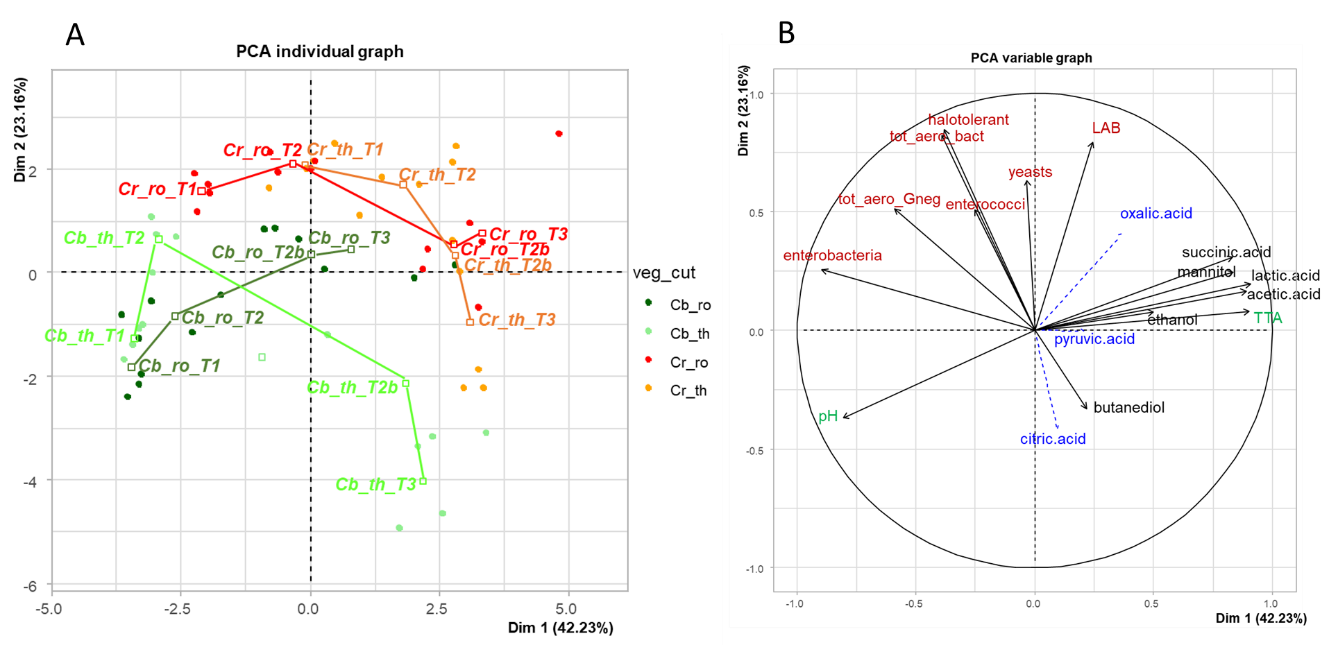
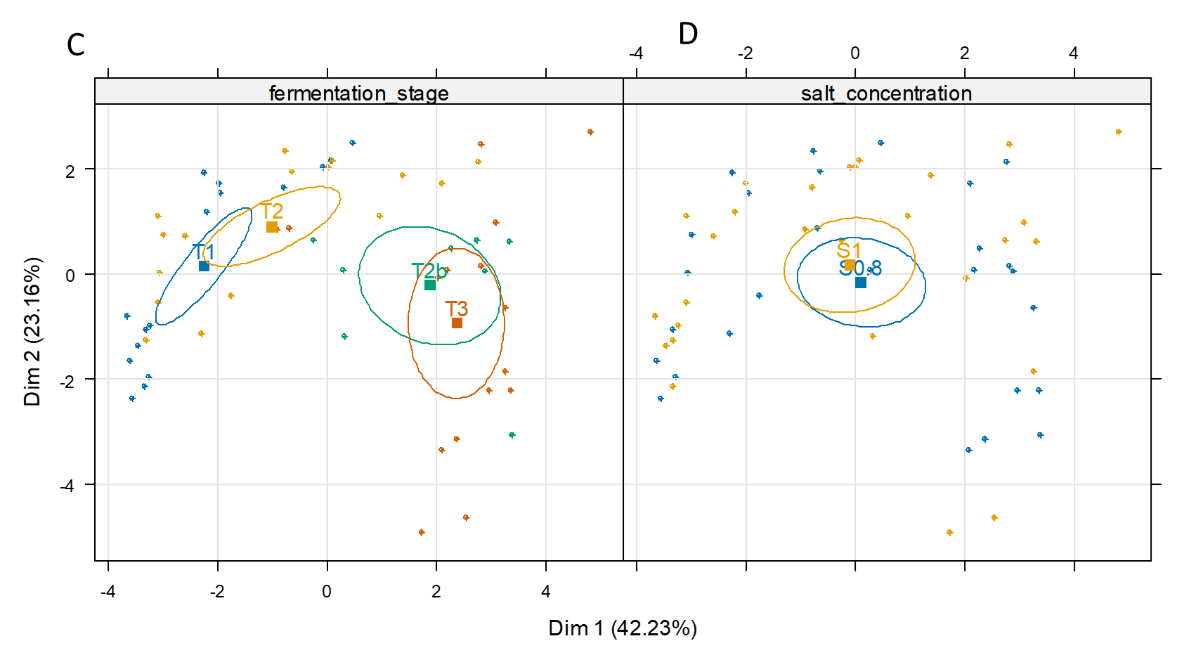
Figure 5 - Principal component analysis compiled using 15 variables: the data on the enumeration of viable microorganisms (n=7, shown in red), levels of the main metabolites (n=6, shown in black) and pH and total titratable acidity (TTA) (shown in green) for 56 samples (cabbage and carrot) analysed at four fermentation stages: T1, 1.7 day, T2, 2.7 (carrot) or 3.6 day (cabbage); T2b (2 weeks) and T3 (4 weeks). On the PCA individual graph (A), the vegetables and their cutting types are abbreviated as follows: Cb_ro, cabbage, rough cutting, in dark green; Cb_th, cabbage, thin cutting, in green; Cr_ro, carrot, rough cutting, in red; Cr_th, carrot, thin cutting, in orange. (B), PCA variable graph: three minor metabolites were not used to calculate the PCA axes but are projected as supplementary variables (blue, dashed lines). The seven targeted microbial groups, are: lactic acid bacteria (LAB), total aerotolerant bacteria (tot_aero_bact), halotolerant bacteria, aerotolerant Gram-negative bacteria (tot_aero_Gneg), yeasts, bile-tolerant Enterobacteriaceae (enterobacteria), and enterococci. For culture media, see Table S2; The two individual maps (C and D) show the 95% confidence ellipses as a function of fermentation stage (C) and salt concentration (D).
Volatile compound profiles also changed over time and were mainly affected by cutting type.
A total of 78 volatile compounds were identified in cabbage and 52 in carrot, with only 21 volatiles were shared by the two vegetables (Supplementary Tables S5 and S6).
Twenty-three sulphur-containing compounds were found in cabbage, including nine (iso)thiocyanates and four nitriles, 23 esters, eight acids, seven alcohols, seven aldehydes, six ketones, and four other compounds (Supplementary Table S5). The abundance of 24 volatiles varied significantly (p-value<0.01) during fermentation, with 15 that increased (fold-change stage T3/stage T1 >2), and 9 that decreased (fold-change stage T1/stage T3 >2). The cutting factor affected the abundance of 15 volatiles, and the salt concentration affected that of ten volatiles, with some interactions between these factors (Supplementary Table S5). PCA performed to illustrate the global effects of stage, cutting and salt on the volatile profiles, showed that cabbage samples were separated on the first axis (24.1% of total variability) depending on the stage of fermentation, and on the second axis (15.9% of total variability) according to their cutting type and salt concentration (Figure 6A, Supplementary Figure S1). Samples at the start of fermentation showed high abundances of many esters, while one-month aged samples contained more ethanol, butan-1-ol, acetic and butanoic acids, and sulphur-containing compounds that originated from cabbage, such as isothiocyanates and nitriles (Supplementary Figure S1).
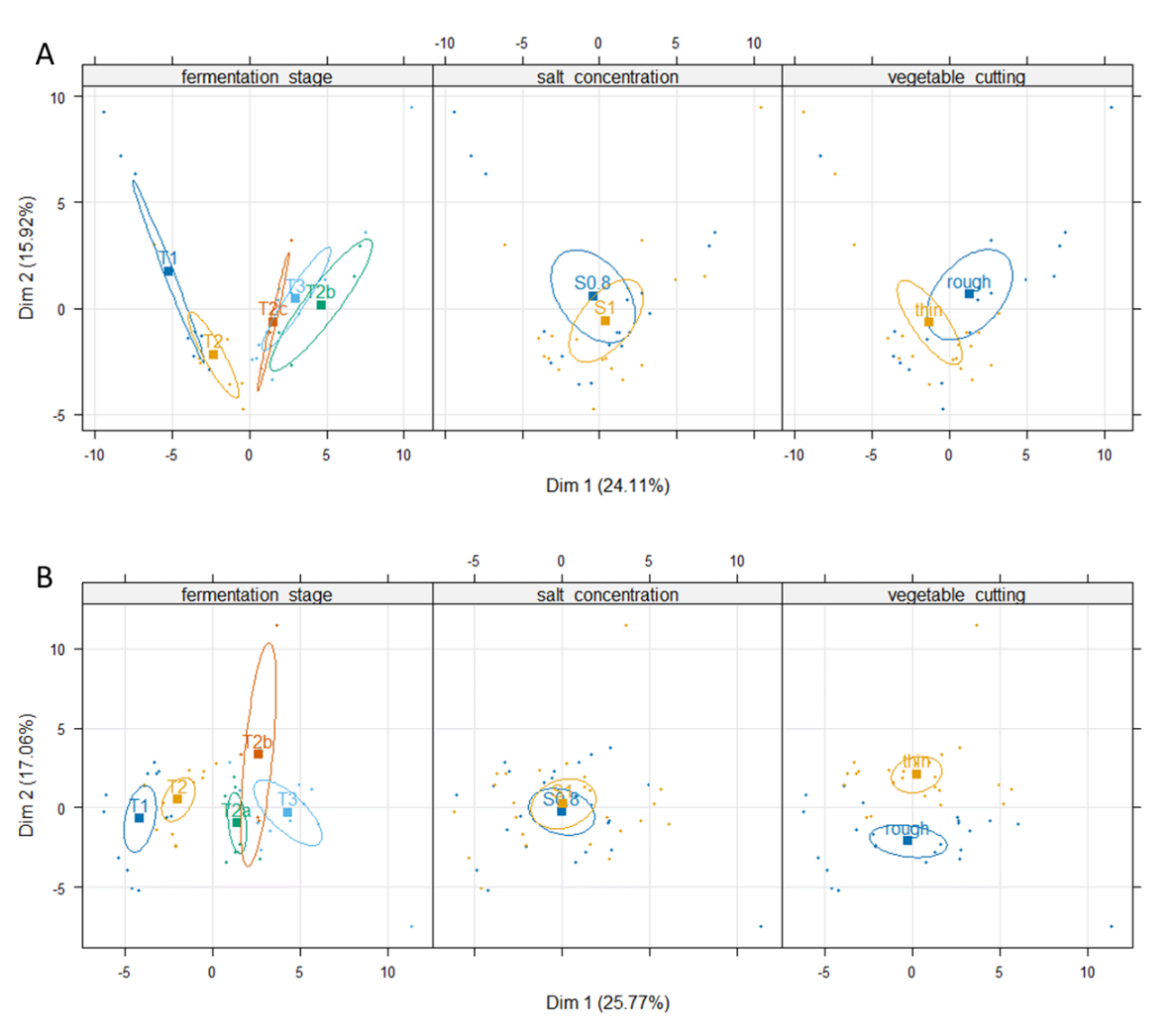
Figure 6 - PCA of volatiles identified in fermented cabbage (A) and fermented carrot (B). The three maps of individuals are coloured and show the 95% confidence ellipses according to the fermentation stage (first panel): T1 (40 h) to T3 (one month), see figure 1 for details on the stages, the salt concentration (second panel): S0.8: 0.8% or S1: 1%, and cutting type (third panel): thin or rough.
Carrot juices included nine ketones, nine aldehydes, eight alcohols, seven esters, five acids, eight terpenes, two sulphur-containing compounds and four other compounds. In addition, many other terpenes and terpenoids were tentatively identified, e.g. γ-terpinene, terpinolene, 1,3,8-p-menthatriene, α-bergamotene, caryophyllene, terpinen-4-ol, β-cyclocitral, (E)-γ-bisabolene, zingiberene, cis-β-farnesene, β-curcumene and β-sesquiphellandrene. These compounds arising from raw carrot were not further considered in this study. The abundance of 35 volatiles varied significantly (p-value<0.01) during fermentation, with 19 that increased in concentration (p-value< 0.01; fold change T3/T0 >2), and nine volatiles that decreased in concentration over time (p-value< 0.01; fold change T0/T3 >2), four of them being aldehydes. (Supplementary Table S6). The compound with the highest fold-change was ethyl lactate (>6000). Cutting type also significantly impacted (p-value<0.01, fold-change >2) the levels of six volatile compounds, which were all more abundant in grated carrot than in sliced carrot. The greatest differences were observed for two terpenes, beta-myrcene and D-limonene, suggesting that grating facilitated their release into brine compared to slicing (Supplementary Table S6). The salt concentration did not affect the levels of volatiles. PCA showed that, as observed in fermented cabbage, the samples were first separated according to their stage of fermentation on the first axis (25.8% of total variability), and on the second axis (17.1% of total variability) according to cutting type, while they not differentiated depending on their salt concentration on the two first axes (Figure 6B, Supplementary Figure S1). Aged samples (stages T2a to T3) contained many volatiles (e.g. ethyl lactate, benzaldehyde, dimethyl trisulphide, esters) while thinly-cut samples were associated with a high abundance of several terpenes (Supplementary Figure S1).
Dynamic changes to juice mineral concentrations and small variations in vitamin concentrations
NaCl concentrations in juices varied significantly depending on the amount of added salt (p-value <0.001), with, on average, 9.06 g/l and 7.08 g/l in 1%-salted and 0.8%-salted samples, respectively. The differences between the salt concentrations observed in juices and the targeted concentrations (0.8 and 1%) resulted from the composition of the coarse grey salt used to prepare the brines. Our results were consistent with the composition of coarse grey salt (e.g. 34 +/- 3 g Na per 100 g product, i.e. 86.4 g NaCl per 100 g product). The differences between observed and targeted salt concentrations could also result from Na migration from the brine to the vegetables during incubation, thus decreasing its concentration in the juice. Significant concentrations of Mg were also supplied to the initial brine by the coarse salt that contained 1 g Mg per 78 g Na. By comparing the total concentrations in juice and the concentration supplied by the added salt, we calculated that the proportion of Mg arising from salt accounted for more than half of the total Mg content in juices (59%, and 64% in thinly- and roughly-cut carrot, respectively, and 51% and 59% in thinly- and roughly-cut cabbage, respectively). We were thus able to calculate the Mg concentration originating from the vegetables alone, referred to as ‘Mg_veg’. By contrast, the coarse salt used did not supply any P and K, and their proportions arising from the salt in juice were negligible (<0.01% and <0.09%, respectively).
The mineral contents in P, K, and Mg_veg (Mg from the vegetable, see above) were 1793, 146, and 27 mg/L, respectively, in carrot, and 1068, 78 and 34 mg/L, respectively, in cabbage (Supplementary Table S7). During fermentation, the concentrations of P and Mg_veg (Mg arising from the vegetable, see above) rose significantly (p-value <0.01) in both cabbage and carrot juice, showing a gradual migration of minerals from the vegetable tissue to the juice, with 32% more P and 52% more Mg_veg after a 1-month incubation (T3) compared to the start of fermentation (stages T1-T2). Cutting type significantly affected (p-value <0.001)the contents in P, K, and Mg_veg of juices. In both vegetables, higher concentrations were observed in the juice of thinly-cut vegetables, with a greater difference in cabbage than in carrot. Juices from shredded cabbage contained, on average, 32%, 18% and 20% more P, K, and Mg_veg, respectively, compared to juices from leaf cabbage, while juices from grated carrot contained 16%, 10% and 13% more P, K, and Mg_veg, respectively, than those of sliced carrot. P and K contents were also globally slightly – but borderline significantly (p-value <0.06) – higher in the more salted samples, with on average, 16% more P and 9% more K in 1% salted cabbage juice, and 8% more P and 5% more K in 1% salted carrot. Trace amounts of Se were also detected in carrot only, at concentrations <0.040 mg/L.
Table 1 - concentrations of vitamins C, B9, B12, K1, K2, of beta-carotene and content in fresh drained vegetables before (T0) and after one month of fermentation (T3). Values are mean and standard deviation of duplicate analyses (except for vitamins K1, K2 and B12 for which only one analysis was performed). nd: not determined; Values that share the same grouping symbol do not significantly differ.
Sample | Code | vit C | vit B9 | β-carotene | vit K1 | vit K2 | vit B12 | dry matter | |
|---|---|---|---|---|---|---|---|---|---|
mg/100 g | µg/100g | mg/100g | µg/100g | mg/100g | µg/100g | % | |||
cabbage T0 | 300-T0 | 10.2c ± 0.39 | 16.5a ± 2.12 | nd | 3.9 | nd | nd | 10.3 ± 0.2 | |
cabbage T3 thin salt 0.8% | 301-T3 | 19.1b ± 1.01 | 14.0ab ± 2.83 | nd | <3.0 | <5 | <0.1 | 18.3 ± 1.2 | |
cabbage T3 thin salt 1% | 311-T3 | 20.4b ± 0.67 | 17.0a ± 1.41 | nd | 3.4 | <5 | <0.1 | 17.0 ± 1.6 | |
cabbage T3 rough salt 0.8% | 321-T3 | 22.2ab ± 0.53 | 11.5abc ± 0.71 | nd | 4.3 | <5 | <0.1 | 17.6 ± 0.8 | |
cabbage T3 rough salt 1% | 331-T3 | 24.8a ± 2.63 | 17.5a ± 0.71 | nd | 6.7 | <5 | <0.1 | 17.7 ± 1.9 | |
carrot T0 | 400-T0 | 1.0d ± 0.11 | 7.65cd ± 0.07 | 11.3a ± 0.3 | <3.0 | nd | nd | 9.2 ± 0.1 | |
carrot T3 thin salt 0.8% | 401-T3 | 1.3d ± 0.22 | 2.35de ± 0.07 | 8.0a ± 2.4 | <3.0 | <5 | <0.1 | 12.6 ± 0.5 | |
carrot T3 thin salt 1% | 411-T3 | 1.2d ± 0.16 | 8.65bc ± 1.77 | 9.8a ± 1.5 | <3.0 | <5 | <0.1 | 12.9 ± 0.1 | |
carrot T3 rough salt 0.8% | 421-T3 | 2.0d ± 0.02 | 1.35e ± 0.21 | 9.3a ± 1.5 | <3.0 | <5 | <0.1 | 13.8 ± 0.1 | |
carrot T3 rough salt 1% | 431-T3 | 1.5d ± 0.02 | 1.65de ± 0.07 | 9.4a ± 1.3 | <3.0 | <5 | <0.1 | 12.6 ± 0.3 | |
The levels of the six vitamins analysed, expressed in mg or µg per 100 g fresh drained vegetable, and the contents in dry matter, are given in Table 1. Compared to raw cabbage, fermented cabbage contained about twice more vitamin C after 1 month, while in carrot, a slight but not statistically significant increase in vitamin C was observed. By contrast, vitamin B9 and beta-carotene tended (p-value = 0.010) to decrease over time. Regarding vitamins B12 and K2, which are not present in the raw vegetables, they were not produced, or the levels were too low to be detected in the fermented vegetables. As for the impact of cutting size, roughly-cut vegetables contained slightly but significantly (p-value<0.03) more vitamin C than thinly-cut ones (+19% and +43% in cabbage and carrot, respectively). In terms of salt concentrations, the vitamin C content was not significantly affected by salt, and vitamin B9 fell more in the 0.8%-salted than in the 1%-salted samples.
Metataxonomic results revealed a rapid replacement of the initial microbiota by enterobacteria and then lactic acid bacteria
The read numbers of sequenced samples ranged from 2,617 to 186,755, with a median value of 29,203 and a mean of 43,484. All samples sequenced after T0 had more than 10,000 reads. A total of 314 ASVs were obtained after 16S rRNA gene sequencing and 2,640 ASVs after gyrB sequencing, each with an abundance exceeding 0.005% of the total. Overall, ASVs belonged to 98 different genera, 62 of which were detected using gyrB only, 12 using 16S only, and 24 common to both markers. The ASVs derived from gyrB sequencing enabled higher taxonomic resolution, with 132 species identified.
The two markers were used in parallel to define the taxonomic profile of the samples at the genus level (Figure 7).
At T0, Pseudomonas largely dominated the bacterial community of both carrot and cabbage samples. The taxonomic profiles then shifted, and Enterobacteriaceae became predominant (Supplementary Figure S3). Some genera were preferentially observed in shredded cabbage (Aeromonas), leaf cabbage (Buttiauxella), or carrot (Erwinia) samples, while others were present regardless of the vegetable (Rahnella, Enterobacter, Serratia). LAB appeared from T1 (Leuconostoc, Lactococcus) with Lactiplantibacillus or Levilactobacillus dominant at T3 in grated and sliced carrot (except for sample 411-a-T3). In cabbage samples, the profile differed markedly, depending on the cutting type. Leuconostoc and Latilactobacillus were detected at T2 in shredded cabbage, while Clostridium developed and became the main genus in leaf cabbage (even if Leuconostoc was detected at T2). Therefore, the taxonomic profiles differed according to the vegetable and cutting type, especially for cabbage, but they did not appear to be impacted to the same degree by the salt concentration.
The taxonomic profiles also varied according to the marker used (Figure 7, Supplementary Figures S3 and S4). Some genera were detected similarly by both 16S and gyrB markers (e.g., Lactococcus, Aeromonas, Erwinia, Pseudomonas), while others exhibited differential detection patterns according to the marker. For example, the Leuconostoc genus was almost undetectable with the 16S marker, while it was detected at a high abundance with the gyrB marker. Conversely, the Clostridium genus was detected with the 16S marker but was scarcely detectable with gyrB marker.
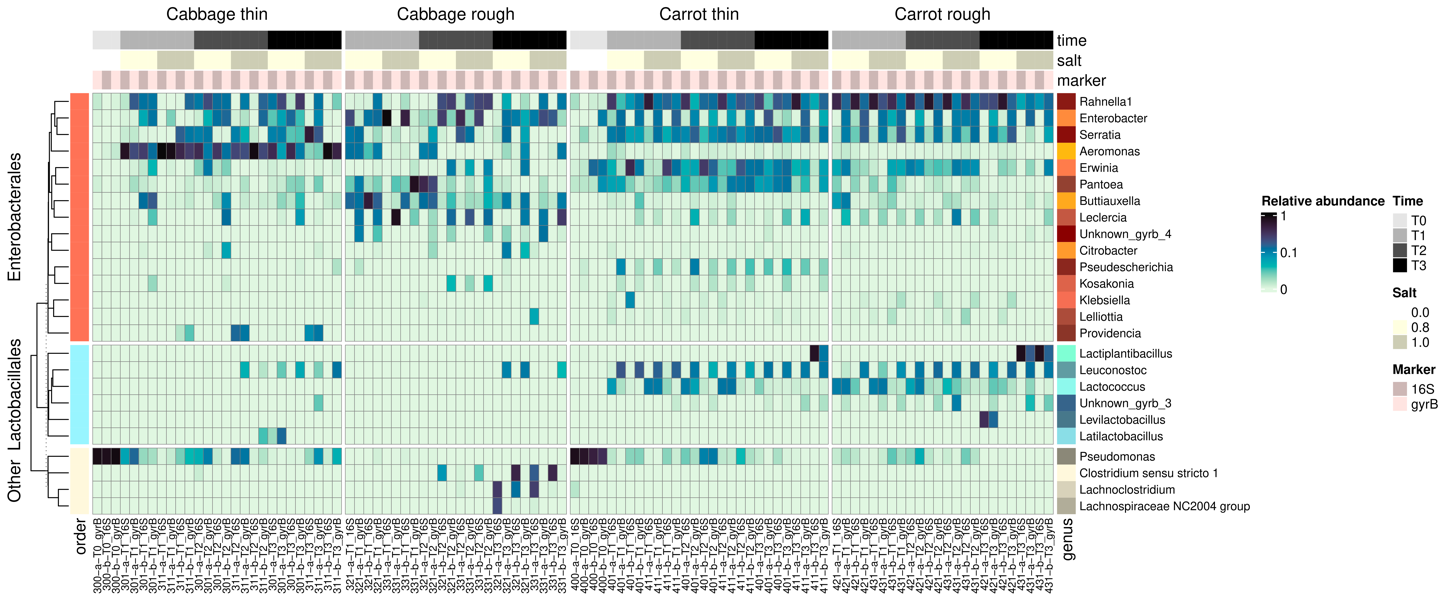
Figure 7 - Heatmaps showing the relative abundance of the 25 most frequently observed genera using both 16S rRNA and gyrB markers. Taxa are coloured (Lactobacillales shown in different shades of blue, Enterobacterales in different shades of orange, and “other” in different shades of grey) and split according to their taxonomic order (Lactobacillales, Enterobacterales and “other”) and then clustered based on the Bray-Curtis dissimilarity. Samples are organised by vegetable and cutting types. Stage and salt level are indicated by an annotation alongside the heatmap.
The higher resolution observed with gyrB compared to 16S made it possible to refine the taxonomic profiles by identifying species (Figure 7), notably those relating to Lactobacillales and Enterobacterales. Regarding Enterobacterales, the species Rahnella aquatilis, Enterobacter sp. 638, Serratia sp. Leaf51 and Pantoea agglomerans were found in both cabbage and carrot samples. For LAB, Leuconostoc mesenteroides, Lactococcus piscium, and Leuconostoc gelidum were present in carrot samples from T1 onwards. At T3, Lactiplantibacillus plantarum and Levilactobacillus brevis were mainly observed. For shredded cabbage samples, L. mesenteroides and Leuconostoc carnosum were present from T2 onwards. L. carnosum and Enterococcus faecalis were observed for leaf cabbage samples at T3. Therefore, taxonomic profiles at the species level also differed according to vegetable and cutting type, but they did not seem to be impacted by the salt concentration. Furthermore, it is worth noting that there were pronounced differences in taxonomic profiles between the duplicate jars (coded a and b). For example, the abundance of the genus Leuconostoc detected with the gyrB marker was much higher in carrot at T1 in replicate 421-b-T1 than in 421-a-T1; the abundance of the genus Lactiplantibacillus detected with the 16S marker was much higher in carrot in replicate 411-b-T3 than in 411-a-T3. In the case of cabbage, differences were particularly marked in roughly-cut cabbage. For example, the abundance of the genus Enterobacter detected with the gyrB marker was much higher in replicate 331-a-T3 than in 331-b-T3. Among the 17 Lactobacillales species identified by gyrB sequencing, 10 were common to the bacterial strains isolated: L. mesenteroides, Lactiplantibacillus pentosus, Enterococcus faecium, L. brevis, L. carnosum, Latilactobacillus curvatus, E. faecalis, L. plantarum, Lactococcus lactis, and Enterococcus casseliflavus. For instance, strains of L. curvatus were isolated from samples 311-b-T2, 331-a-T2 and 301-a-T3 and were effectively detected with gyrB sequencing in those samples.
In earlier sections of this paper, it was shown that the fermentation rate varied by cutting type and, to a lesser extent, by salt concentration. Additionally, it was important to investigate whether the fermentation profiles still differed or eventually converged at one month. A multi-block Partial Least Squares-Discriminant Analysis (PLS-DA) was therefore performed to determine whether the samples exhibited different signatures after one month of fermentation (stage T3), given their cutting type and/or salt concentration. The datasets included gyrB and 16S metataxonomic profiles, levels of the five main metabolites, viable counts for six bacterial groups and yeasts, and some selected volatile compounds, selected because they result from different synthesis pathways and displayed marked changes in abundance during fermentation. Models based on cutting type only enabled the identification of a discriminant signature among the samples, unlike models based solely on salt concentration or on both salt concentration and cutting type. The results of the PLS-DA on carrot and cabbage samples are shown in Figure 8. The model discriminated the samples based on cutting type according to variables belonging to the different datasets (Figure 8).
Regarding cabbage, the first dimension distinguished the samples based on cutting type across all datasets (p-value <0.05 for all datasets, Wilcoxon test). Thinly-cut samples exhibited a relatively homogeneous signature, characterised by high concentrations of acetic acid, lactic acid and mannitol, a low pH, a higher abundance of Leuconostoc, and overall lower bacterial viable counts. Roughly-cut cabbage samples showed a signature marked by high ethanol concentrations, the presence of Clostridium, Lachnoclostridium, Lachnospiraceae, and a higher abundance of Buttiauxella. One of the replicates of roughly-cut cabbage, 331-a-T3, was a particular case with a higher abundance of Enterococcus and Enterobacter, higher levels of acetoin and diacetyl and lower levels of the main metabolites (mannitol, acetic and lactic acids) compared to the three other roughly-cut cabbage samples
For carrot, the first dimension distinguished samples based on cutting type only for the gyrB and metabolite datasets, while the 16S, enumeration and volatile datasets failed to discriminate the cutting type of carrot samples. Thinly-cut carrot samples had a signature characterised by a higher abundance of Pseudescherichia, while roughly-cut carrot samples were characterised by a higher abundance of Buttiauxella, higher levels of lactic acid, ethanol and butanediol, and a lower pH. Fermentation profiles differed between carrot samples independently of their cutting type. For example, one thinly-cut sample, 411-b-T3, displayed a profile similar to that of two roughly cut samples (431-a-T3 and 431-b-T3) characterised by a lower pH, a higher abundance of Lactiplantibacillus and higher levels of lactic acid, ethanol, acetoin and diacetyl. Therefore, carrot profiles did not exhibit a strong discriminating signature by cutting type across the different blocks (except for gyrB and metabolites), as a convergence between samples with different cutting types occurred at one month.
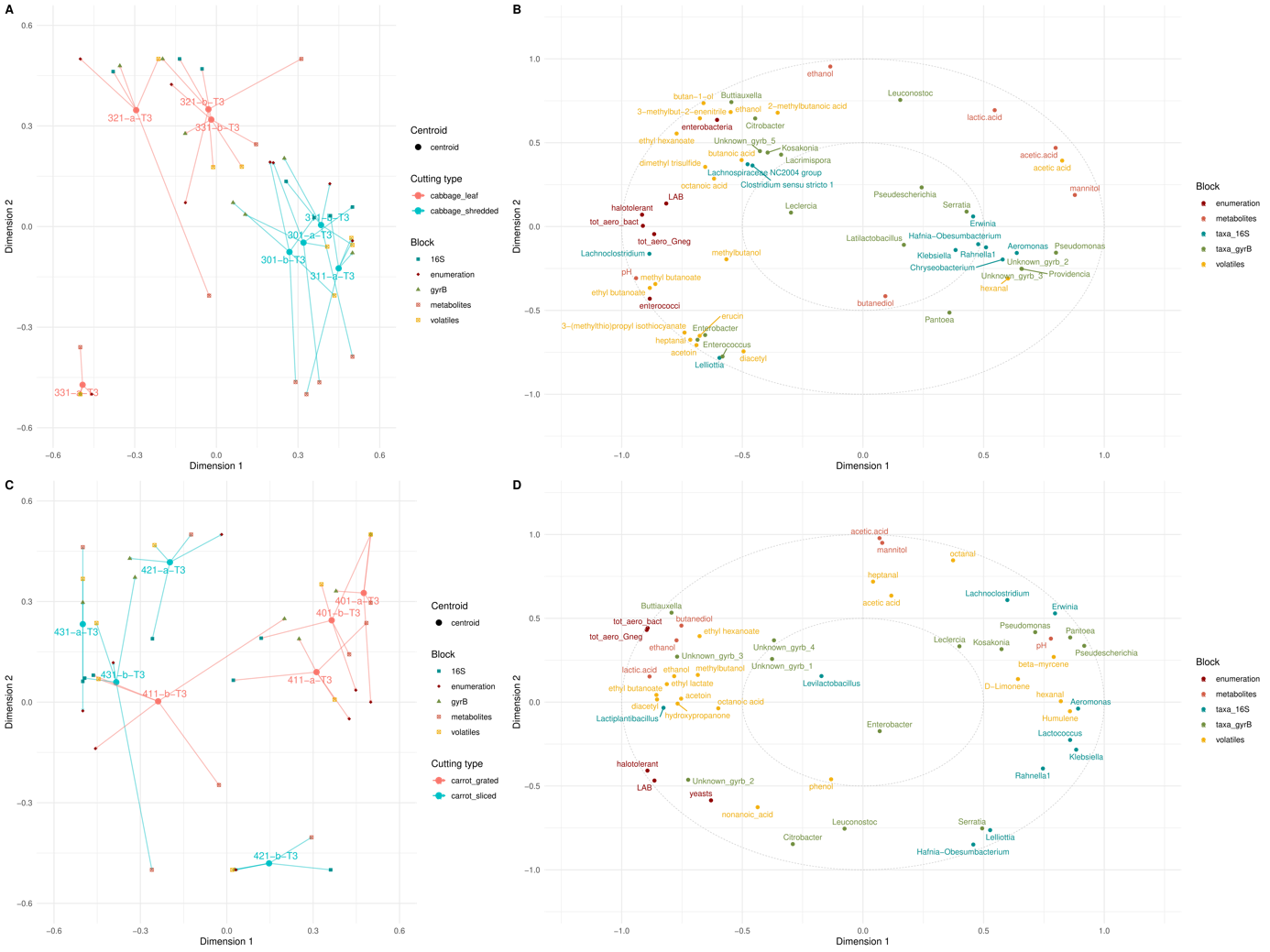
Figure 8 - Multiblock PLS-DA results for cabbage (A, B) and carrot (C, D) samples at stage T3 (one-month fermentation). The left-hand panels (A, C) show the alignment of samples in the latent space, where each round point represents the centroid of all datasets for a given sample, and the arrow tips indicate the sample's position within each block. The blocks are colour-coded as follows: blue for 16S rRNA gene taxonomic data, red for microbial counts, green for gyrB taxonomic data, orange for metabolites, and yellow for volatiles. The right-hand panels (B, D) present correlation circle plots showing the relationships between variables as scatter plots, with variables coloured according to their respective blocks. The microbial count variables correspond to seven targeted microbial groups: lactic acid bacteria (LAB), total aerotolerant bacteria (tot_aero_bact), halotolerant bacteria, aerotolerant Gram-negative bacteria (tot_aero_Gneg), yeasts, bile-tolerant Enterobacteriaceae (enterobacteria), and enterococci.
Discussion
The experimental design allowed us to highlight a role for vegetable cutting despite considerable jar-to-jar variability
The objective of the present study was to better understand the microbial dynamics and biochemical changes resulting from the spontaneous fermentation of vegetables by combining several omics approaches: culturomics, 16S rRNA gene and gyrB metataxonomics, and targeted metabolomics. More specifically, our aim was to investigate the effects of two factors, cutting type and a 20% reduction in the amount of salt added, on microbial and biochemical changes during fermentation. We chose both a root and a leafy vegetable, carrot and cabbage, which are commonly used to make fermented vegetables in France (Thierry et al., 2023b). The main differences between the two vegetables used in our study concerned the rate of fermentation and the final concentrations of metabolites. However, the experiment was not designed to compare carrot and cabbage directly, because the vegetable cultivar, cultivation conditions, harvest and storage conditions (time, temperature) can also impact their microbial and biochemical composition and hence their fermentation (Leff & Fierer, 2013). Moreover, the processing steps e.g. the washing of carrot vs only the removal of external cabbage leaves, could also induce differences. As a result, the differences observed during the present study between carrot and cabbage fermentation should not be generalised. The impact of cutting type or size has only rarely been addressed, although this factor may markedly vary, at least in a domestic setting (Thierry et al., 2023b). Regarding the NaCl content in foods, the World Health Organisation has suggested reducing sodium intake by 30% to achieve the WHO guideline of 2 g day−1 (i.e., 5 g salt day−1) by 2025 (World Health Organization, Regional Office for Europe, 2018). We chose to compare a salt concentration of 1% (the level generally recommended) with one of 0.8%, i.e. a 20% Na reduction. Only limited differences in microbial and biochemical changes were observed, in carrot only, between these two salt concentrations, although mineral diffusion was slightly enhanced at the higher concentration (see below). Considerable jar-to-jar variability was observed, which may have limited any demonstration of the effect of salt reduction. Each jar exhibited its own fermentation path and showed some specificity regarding the microbial results, metabolite profiles and acidification rate (e.g. illustrated on PCA in Figure 5). Each jar was prepared by taking the required amount of cut vegetables from a large (~10 kg) bin, without previously mixing the bin’s content. We therefore hypothesise differences in the initial microbiota present in each jar, particularly concerning LAB, which were highly sub-dominant or even below the detection threshold in raw vegetables. Differences could also result from the composition of different carrots and different cabbages. The variations we observed were both quantitative (i.e. differences in the rate of fermentation) and qualitative (e.g. the nature of the dominant taxa that grew over time). For example, yeasts were detected in only one of the duplicate jars in different samples (leaf cabbage at 40 h fermentation, sliced carrot at one month of fermentation). Variability between replicate jars was also evidenced from the metabolite profiles (Figure 3) and metataxonomic profiles (Figure 7). Heterogeneity can occur in the tanks used for industrial sauerkraut production, so the juice is recirculated to prevent this source of potential defects (Pederson & Albury, 1969). Differences between replicates had also been observed, but were not discussed, in several other studies, e.g. the results regarding 16S-based metataxonomics in fermented radish and carrot (Raghuvanshi et al., 2019), and in paocai (Wang et al., 2020; Wang et al., 2022a). For example, the abundance of Enterobacteriaceae according to the 16S rRNA gene metataxonomics ranged from ~20% to ~80% in the triplicates of four-day fermented carrot and one-day radish, and high variations in pH during the first two days of fermentation were also observed (Raghuvanshi et al., 2019). These results stress the need to include a sufficient number of replicates in studies on spontaneous vegetable fermentation, particularly at a small scale.
LAB quickly outcompeted bile-tolerant Enterobacteriaceae that dominated the plant microbiota at the beginning of fermentation
Both culturomic and metataxonomic results on both vegetables confirmed the sequential establishment of the microbial community, with the presence of a large proportion of Pseudomonas in raw vegetables and the early development of bile-tolerant Enterobacteriaceae, referred to as enterobacteria below. The dominance of Pseudomonas and enterobacteria during the first hours of fermentation was consistent with the fact that these two groups constitute a large part of plant surface microbiota (Lund, 1992; Leff & Fierer, 2013; Jackson et al., 2015). Pseudomonas members are characterised by their great ability to colonise different ecological niches and have been recovered from a wide variety of 77 samples from 11 different vegetables (Ruiz-Roldán et al., 2021). In the present study, fewer viable enterobacteria were found in raw carrot than in raw cabbage (4.3 and 6.0 logCFU/g, respectively) which may have been due to the fact that the carrots were washed before use, unlike the cabbages. Enterobacteria are largely represented in the microbiota of fresh vegetables, e.g. at ~5.2 logCFU/g in 41 out of the 45 samples characterised, which included carrot, cabbage, and five other fresh vegetables (Al-Kharousi et al., 2016). The first shift conserved among different fermented vegetables was from the initial microbial population of vegetables to Enterobacterales, according to a recent integrative bioinformatics approach used to perform a meta-analysis of 10 public amplicon data sets on fermented vegetables (Junker et al., 2024). For example, members of the Enterobacteriaceae family that included many non-affiliated taxa, and Erwinia, largely dominated on different vegetables, according to a 16S-based metataxonomic analysis (Raghuvanshi et al., 2019).
Our results also confirmed the early development of LAB, which managed to outcompete Pseudomonas and enterobacteria notwithstanding their prevalence in raw vegetables. This was fully in line with the results of a bioinformatics meta-analysis on different fermented vegetables which showed that after Enterobacterales domination, a second shift led to an assemblage dominated by Lactobacillales, i.e. LAB (Junker et al., 2024). This second shift has been observed, for example, in carrot juice (Wuyts et al., 2018), cucumber (Stoll et al., 2020), kimchi (Song et al., 2020; Jung et al., 2022), and paocai (Wang et al., 2020). LAB are able to live as endophytes in a wide variety of crop plants (Pontonio et al., 2018). In our study, viable LAB were non-detectable in cabbage and their initial counts were 3 logCFU/g in carrot, in line with the values of 2 to 4 CFU/g previously reported (Di Cagno et al., 2013). Plant endophytic communities are dominated by Gammaproteobacteria, Alphaproteobacteria and Actinobacteria. Firmicutes are sub-dominant and mainly represented by Bacillales, while Lactobacillales are generally under 0.1% of relative abundance (Hacquard et al., 2015; Kõiv et al., 2019). LAB are often not detected in starting ingredients by metatoxonomics, as has been shown in sauerkraut manufacture (Zabat et al., 2018). The selective pressure of the environmental conditions that result from the fermentation process applied, and particularly the anaerobic conditions, indeed favours the growth of LAB to the detriment of enterobacteria (Yu et al., 2020). LAB growth leads to significant acidification of the environment, thus inhibiting the growth of enterobacteria that cannot withstand the acidic pH (Ostling & Lindgren, 1993). Concomitantly with acidification, we effectively observed a drastic reduction in viable enterobacteria, which were no longer detected by plate counting after two weeks in three of the four conditions studied, i.e. in shredded cabbage and both thinly and roughly-cut carrot, which were characterised by pH <4.0 and titratable acidity >0.45%. These results are in line with the safety threshold recommended by the Codex Alimentarius standard for pickled fruits and vegetables, which stipulates that the product must be prepared and packaged “to ensure an equilibrium pH of less than 4.6” (FAO, 2007). Our results further illustrate the ubiquitous nature of LAB in the context of food fermentation, thanks to their ability to rapidly ferment different carbohydrates into lactic acid (Gänzle, 2015). As well as the inhibitory activity of the organic acids they produce, LAB may also inhibit enterobacteria through the synthesis of antimicrobial peptides, since many of the species found in our study are known to potentially produce bacteriocins (Zacharof & Lovitt, 2012; Hernández-González et al., 2021). A small proportion of LAB (when compared to that of Enterobacteriaceae) was observed using metataxonomics in the present study, even after one month of fermentation, and particularly in fermented cabbage. In a previous study using the same methods to characterise domestic samples of fermented vegetables, metataxonomic results showed that LAB had a median abundance of 90%, mainly represented by members of the Lactobacillaceae family, with Enterobacterales as the second main taxon (Thierry et al., 2023b). Moreover, a negative relationship was observed between the abundance of Enterobacterales and the age of the samples. The lower LAB abundance observed in the present study could thus be explained by the age of samples: one month maximum in the present study versus a median duration of six months (ranging from two weeks to four years). Similarly, LAB dominated in a study on paocai fermented at 15°C and 25°C for one year, representing about 60% of total abundance, with (former) Lactobacillus as the dominant genus (Wang et al., 2020).
During our study, the first LAB species that grew in both carrot and cabbage were heterofermentative: Leuconostoc sp. and L. lactis, in line with previous reports in other fermented vegetables, such as paocai (Wang et al., 2022b). Heterofermentative LAB are able to degrade a wide variety of carbohydrates (Gänzle, 2015) and Leuconostoc efficiently metabolises sucrose (Cogan & Jordan, 1994).
The choice of metataxonomic markers markedly impacted the picture of the bacterial community. Metataxonomics using both 16S rRNA and gyrB genes as markers helped us to describe changes to the bacterial community during fermentation. The gyrB marker, although rarely used in metataxonomics, offers species-level taxonomic resolution in food ecosystems (Poirier et al., 2018b), as effectively observed in the present study. For the 16S marker, the V3-V4 region is the most commonly used in the field of food microbiology (Parente et al., 2022), but we chose the V5-V7 region to compare the results with that of our previous study on fermented vegetables (Thierry et al., 2023b). Both markers confirmed the succession of Enterobacteriaceae and LAB, except in some roughly-cut cabbage samples. However, the gyrB marker uniquely detected Leuconostoc as the first LAB genus to appear and this was also confirmed by the results of the culture-dependent approach. In three roughly-cut cabbage samples, the 16S marker uniquely detected the Clostridium taxon. Detection of this undesirable genus is important because some Clostridium species are responsible for food poisoning. However, we failed to find any other study that reported clostridia-related poisoning associated with the consumption of fermented vegetables.
Metabolites of fermented products as markers of microbial activity
The compounds analysed during the present study were carbohydrates, organic acids, alcohols and volatile compounds. Mannitol, lactic and acetic acids were the main metabolites produced, in line with previous reports on fermented carrot juice (Wuyts et al., 2018) and sauerkraut (Plengvidhya et al., 2007; Tlais et al., 2022). Mannitol, lactic acid and acetic acid respectively accounted for about 45-55%, 22-30%, and 10-12% of total metabolites in carrot and shredded cabbage after one month of fermentation. These proportions were similar to those reported in carrot juice (Wuyts et al., 2018), sauerkraut (Plengvidhya et al., 2007), and other fermented vegetables as kimchi (Jung et al., 2011). These metabolites are typical markers of heterofermentative LAB metabolism, as the Leuconostoc members identified in both cabbage and carrot convert fructose into mannitol (Wisselink et al., 2002; Martínez-Miranda et al., 2022). They also convert other carbohydrates into lactic acid, acetic acid and ethanol, while homofermentative LAB convert carbohydrates into lactic acid as the main end-product. In our study, the ratios of lactic and acetic acids after one month of fermentation varied from one jar to another, from 1.5 to 5 in cabbage and 1.7 to 4.3 in carrot. Butanediol was also detected in some samples, at markedly variable concentrations. Butanediol can be produced from acetoin by L. mesenteroides and Latilactobacillus sakei. For example, metatranscriptomic analyses in kimchi have shown that genes encoding the pathway from pyruvate to diacetyl/acetoin and butanediol were expressed during kimchi fermentation (Chun et al., 2017; Kim et al., 2020). Concerning volatiles, most of them cannot be used as markers for the activity of specific microbial groups because their pathways of formation are shared by many bacterial groups. For example, isothiocyanates, thiocyanates, and nitriles were detected by GC-MS in fermented cabbage samples (Supplementary Table S5). These sulphur-containing compounds derive from the glucosinolates present in cabbage, which were completely degraded after one month of fermentation (results not shown), as previously reported (Wieczorek & Drabińska, 2022). Even if some LAB strains can hydrolyse glucosinolates into nitriles, other bacteria such as enterobacteria also do so (Mullaney et al., 2013). Moreover, the activity of plant myrosinase also results in glucosinolate hydrolysis, and is favoured by both the cutting of vegetables (which releases glucosinolates and myrosinase from the separated cell parts) and by the fall in pH, which favours myrosinase activity (Wieczorek & Drabińska, 2022). As for biogenic amines, which are mainly produced by Enterobacteriaceae from amino acid decarboxylation (Halász et al., 1994), they were not analysed during our study. Total concentrations of biogenic amines can reach from ~50 to ~600 mg/kg in different fermented vegetables (Świder et al., 2020).
Thin cutting favours the release of solutes and increases the fermentation rate
The cutting of vegetables before fermentation ranges from thin shredding to large pieces in domestic production (as stated above), but to the best of our knowledge, the effects of cutting have been little studied. A “degree of disintegration” was referred to, alongside temperature and type of vegetable, among the factors influencing fermentation (Buckenhuskes, 1993; Buckenhueskes, 2015). The nutrients present inside vegetable cells need to be released in the aqueous phase, i.e. juice that is made available to the microorganisms via shredding, slicing or just piercing, depending on the vegetables concerned (Buckenhueskes, 2015). The thinner the cutting, the greater the surface area of cut plant tissue that can directly release vegetable solutes into brine, thus supplying LAB with nutrients and increasing the buffering capacity of the brine. Solutes can also diffuse from entire, undamaged vegetables, as described in cucumber, but at a lower rate (Passos et al., 2005). In a study designed to model the equilibrium of solutes between brine and entire cucumber (peeled or not), it was shown that the diffusion coefficient of glucose was 9.2 times higher for peeled than for unpeeled cucumber (Potts et al., 1986), which suggests that the surface of cut tissue is important to promoting the diffusion of solutes into brine. We therefore calculated the surface of cut vegetables by estimating the mean dimension of the pieces used in the present study, assimilated them either to cylinders (entire, sliced and grated carrot), or to parallelepipeds (cabbage leaf pieces and shredded cabbage). Moreover, the experimental mass of vegetable and brine weighed per jar differed depending on the vegetable and the cutting method, thus resulting in differences in the ratio of vegetable to brine, which ranged from 0.73 (leaf cabbage), 0.94 (shredded cabbage), 1.24 (sliced carrot) to 1.33 (grated carrot). Based on these ratios and the cut tissue surface estimates, we calculated the cut surfaces per g of initial brine, which were approximately 0.4 cm2, 8 cm2, 9 cm2, and 19 cm2, for leaf cabbage, sliced carrot, shredded cabbage, and grated carrot, respectively, as detailed in Supplementary Table S8. In other words, the cut surface of the thinly-cut cabbage was approximately 26-fold higher than that of roughly-cut cabbage, while it was only 2-fold for the thinly-cut carrot compared to roughly-cut carrot. We therefore hypothesised that these marked differences in cut surface could, at least partly, explain why we found that the two cutting types of cabbage differed markedly in terms of the acidification and viable enterobacteria decrease rates, in contrast to the smaller differences observed between sliced and grated carrots (Figure 2).
The hypothesis of a greater diffusion of vegetable solutes into brine with thinly-cut vegetables was further supported by our results regarding the mineral composition of juices. First, they contained 18-32% more minerals (K, P, and Mg) arising from the vegetable tissue in shredded than in leaf cabbage, and 10-16% more in grated than in sliced carrot. Secondly, carrot juices contained more P and K than cabbage juice, although white cabbage was expected to be as rich in K and 1.5 richer in P compared to carrot, according to Ciqual data (Anses, 2020), thus suggesting a higher global diffusion from vegetable into juice in the case of carrot, for which the cut surface was greater. As a result, we can hypothesise that the buffering capacity of the juice was higher for thinly-cut vegetables than for roughly-cut ones. This assumption is consistent with the fact that, for a given pH, TTA was higher in thinly-cut vegetables. For example, TTA was twice as high in grated carrots at T1 as in sliced carrots, despite a similar pH. The cutting effect had previously been studied in order to compare the survival of pathogenic strains during the fermentation of cabbage as whole heads or shredded (Niksic et al., 2005). These authors attributed the lower survival observed with shredded cabbage to the significantly higher total titratable acidity in shredded cabbage juice compared to that of whole head cabbage, linked to the higher buffering capacity of the juice. The importance of the buffer capacity of vegetable juice has previously been highlighted. A pioneering study showed that the composition of cucumbers depended on their size. Smaller cucumbers contained lower levels of sugars and a higher natural buffering capacity than larger ones and achieved complete sugar utilisation during fermentation (Lu et al., 2002). Buffer models were later developed in cucumber juice of different compositions, so as to be able to assess the relationship between pH and the concentration of acids (Breidt & Skinner, 2022).
The addition of salt is known to draw nutrients from the vegetable tissue towards the juice. And indeed, during the present study, we saw a slight but significant effect of the amount of added NaCl on the release of P and K in the juice (+ 5-8% in carrot, and + 9-16% in cabbage).
In the case of leaf cabbage, which had by far the smallest cut surface, we also observed some cases of faulty fermentation. For example, two of the four jars of leaf cabbage characterised still contained live enterobacteria after one month of fermentation (321-a and 331-b); the third (321-b) contained lower viable LAB counts (7.6 logCFU/mL) and a high content in ethanol (5.2 g/mL vs 1.6 g/mL in all other samples at that stage); and the fourth (331-a) contained enterococci as the only LAB species, and was associated with an atypical metabolite profile without mannitol (Figures 2, 3, and 4) and displayed a distinct volatile profile, with the highest levels of butanoic acid esters, for example. The four leaf cabbage samples also contained Clostridium (321b, 331a, 331b) and/or Lachnoclostridium (321a, 321b, 331a) taxa. The mean pH of these four samples was 4.0, versus 3.6 at the same stage in all other samples. It is worth mentioning that many leaf cabbage samples had a very unpleasant and atypical smell. We therefore hypothesise that the cutting of cabbage into large leaf pieces was insufficient to supply LAB with sufficient nutrients and that there may be a threshold cut surface below which rapid lactic fermentation is hardly achieved.
In brief, fine cutting, as well as salting, favours the release of solutes towards the juice, thus supplying microorganisms with the nutrients they require to grow and increasing the buffer capacity of brine. Consequently, more rapid acidification, a higher titratable acidity and a more rapid fall in the number of viable enterobacteria can be expected, as was effectively observed in the present study and particularly for cabbage, where the surface generated by the cutting step was much greater (> 20-fold) in shredded cabbage than in leaf cabbage. The difference in salt concentration also intensified solute release, as observed in the present study for some minerals, but the possible effects on microbial growth were probably masked by the jar-to-jar variability observed. In line with the importance of the buffer capacity, the targeted values for both pH (< 4) and titratable acidity (1%) are shown in the specifications for the protected geographical indication of “Sauerkraut of Alsace”, in order to satisfy safety and sensory requirements (EU Commission implementing regulation, 2018). It should also be underlined that for sauerkraut manufacture, shredded cabbage is generally salted using dry salt and not brine (as performed during our study) which induces a higher buffering capacity of juice in the former than in the latter.
Some health benefits and risks associated with fermented vegetable consumption
As well as the preservation of vitamins (discussed below), fermented foods have been associated with potential health benefits that result from two main factors. The first is the microbial production of metabolites of interest for human nutrition (e.g. vitamins, bioactive peptides) or those that can positively affect human health (e.g. mannitol, γ-aminobutyric acid) (Lenhart & Chey, 2017). The second is the presence of live microorganisms that can interact with the intestinal microbiota (Rezac et al., 2018). Initially widely explored in the context of dairy-based fermented foods (Kok & Hutkins, 2018; Companys et al., 2020) and Korean kimchi (Cha et al., 2023), this has now also been clearly demonstrated for other non-dairy foods (Wuyts et al., 2020; Valero-Cases et al., 2020).
Conversely, in the case of poorly controlled production, the consumption of altered fermented products can constitute a health risk. In terms of fermented vegetables, the large number of enterobacteria present at the start of fermentation is associated with the risk of toxin and biogenic amine production. The presence of spore-forming bacteria, including toxin producers such as certain Clostridium sp., has also been reported. In a recent quantitative risk assessment study in South Korea regarding Clostridium perfringens foodborne illness via kimchi consumption, the authors concluded that the risk was “very low” (Choi et al., 2020). Enterobacteria disappear as soon as the pH has remained low enough for a period of time, and not all of them constitute hazards. For example, in the present study, we mainly identified Hafnia alvei among the clones isolated on the VRBG medium from the leaf cabbage samples in which enterobacteria were alive after one month of fermentation. H. alvei is commonly isolated from raw milk Camembert cheese and even used as an adjunct culture in this cheese. One of the leaf cabbage samples also contained clostridia. Fortunately, none of these samples would have been consumed because they had a very unpleasant odour.
We focused in this study on the vitamin contents in raw vegetables and after fermentation. Vitamin levels have been shown to fall during fermentation in most reported cases, as stated in a recent literature review (Thierry et al., 2023a). The vitamin C, vitamin K1 and beta-carotene contents in raw carrot and/or cabbage observed in the present study were in line with the values expected from nutritional food tables, while the vitamin B9 content was within the lower range of reported values (Anses, 2020). We only observed an increase in vitamin C during fermentation, which may have been due to the microbial activity of ascorbigen degradation (Berger et al., 2020).
Acknowledgments
We would like to thank Gilles Drege from CTCPA (Agri-food Technical Centre), Nantes, France for his help with organising and preparing the jars of sliced and grated carrot, at the Oniris platform, Nantes, and Patrice de Martrin and Geoffroy Branjonneau, PROTIAL, Beaucouzé, France, for their help with organising and preparing the jars of shredded and leaf cabbage.
We are grateful to staff at the INRAE MIGALE bioinformatics facility (MIGALE, INRAE, 2020. Migale bioinformatics Facility, doi: 10.15454/1.5572390655343293E12) for their assistance with computing and storage resources.
We also thank Cécile Grondin, UMR SPO, INRAE, Université de Montpellier, Institut Agro, Montpellier, France, for yeast isolate identification, Véronique Broussole and Stephanie Oriol, UMR408 SQPOV, INRAE, Montpellier, France for Bacillus spp. identification, and Arlette Leduc, UMR STLO, INRAE, Institut Agro, Rennes, France for her help with mineral analysis.
Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Microbiology (https://doi.org/10.24072/pci.microbiol.100158; Marsit, 2025).
Funding
This work received financial support from the Conseil Régional de Bretagne, France, the Conseil Régional des Pays de la Loire, France, and Fonds de dotation EKIP, Paris, France, in the framework of the FLEGME inter-regional project, under the scientific coordination of INRAE and managed by the VEGEPOLYS VALLEY competitive cluster.
Conflicts of interest disclosure
The authors declare they comply with the PCI rule of having no financial conflicts of interest.
Data, scripts, code, and supplementary information availability
Data and scripts are available online in the data.gouv repository: https://doi.org/10.57745/MJWSJQ (Thierry, 2024). The sequence data are available online in the European Nucleotide Archive (ENA) at EMBL-EBI: PRJEB79032 on the webpage hosting the data https://www.ebi.ac.uk/ena/browser/view/PRJEB79032. The supplementary information are available online in the HAL repository: https://hal.inrae.fr/hal-04701063v2 (Valence et al., 2025).