
 CC-BY 4.0
CC-BY 4.0
Introduction
Islands harbour the most vulnerable ecosystems affected by global change (Fernández-Palacios et al. 2021a). Approximately 75% of extinctions have occurred on islands, and over half of all terrestrial species that face imminent extinction are island-dwellers, with invasive alien species and land-use change as leading drivers of species’ declines. Climate change emerges as a growing threat for island biota and it may interact in unexpected ways with invasive alien species and land-use change (Mantyka-Pringle et al. 2011; Capdevila et al. 2021). As a result, islands are commonly considered the epicentres of past, imminent, and potential future species extinctions (Supplementary Material S1).
Island ecosystems are disproportionately vulnerable to threats for three key reasons. Most insular species are intrinsically more sensitive to any given threat due to their specific traits deriving from the so-called ‘island syndrome’ (Lomolino 1985; Biddick et al. 2019; Benítez-López et al. 2021; Rozzi et al. 2023). Second, insular species are less likely to adapt to new threats due to their inherent demographic features (e.g., small population sizes, naturally fragmented distribution ranges). Lastly, the physiography of islands, specifically in the case of isolated, small-sized ones, renders their biota more exposed to threats and also less able to escape compared to their mainland counterparts (Fernández-Palacios et al., 2021a; see also the “uniqueness of insular biota” section for a more complete description of those inherent vulnerabilities).
Despite the urgency of the need to protect unique and highly vulnerable island biodiversity from ongoing global change, insular biota receives very cursory attention in international biodiversity policy frameworks. In December 2022, the 196 parties of the Convention on Biological Diversity (CBD) adopted the Kunming-Montreal Global Biodiversity Framework (KM-GBF, https://www.cbd.int/doc/decisions/cop-15/cop-15-dec-04-en.pdf), involving 23 action-oriented global targets. Among them, only one target (Target 6) explicitly mentions islands as priority areas for conservation (though not explicitly mentioning insular biota) and two other targets (Target 3 and 4, GBF) require strong actions on islands, reflecting a broader scientific bias toward mainland (Rodrigues et al. 2010). Since the emergence of climate change vulnerability assessments, the vast majority of those assessments have been designed for, and applied to, mainland ecosystems. When extended to islands, these frameworks are often mere adaptations of those developed for mainland systems. This approach is particularly problematic, as it fails to account for the unique complexities and specific characteristics of insular biota. Consequently, our knowledge of the vulnerability of insular ecosystems and species is incomplete and results most likely in an underestimation of their global vulnerability.
Here, our objective is to introduce an adapted framework that addresses the limitations of existing approaches by incorporating the idiosyncrasies of island biota. This framework is specifically designed to quantify the vulnerability of terrestrial insular biota to multiple threats and aims to capture the distinct challenges posed by the unique attributes of insular ecosystems, such as the island syndrome, the isolated nature of islands, and their high levels of endemism (see also Figure 1A). The goal is also to provide improvements of existing vulnerability frameworks that are not specific to island ecosystems (e.g., the inclusion of multiple threats and dimension of diversity). We define vulnerability in this framework across multiple biodiversity dimensions, considering the exposure, sensitivity, and adaptive capacity of insular biota to multiple threats. The originality of our framework is that it is specifically designed for insular biodiversity, with the inclusion of multiple threats, taxa, and dimensions of diversity, such as functional and phylogenetic diversity, as well as the inclusion of vulnerability markers at species, community and assemblage levels of islands. We first describe the characteristics that contribute to the vulnerability of island biota to global changes, which need to be considered in insular biodiversity vulnerability assessments. Then, we describe how our framework can address some of the ongoing questions on island biota vulnerability.
Challenges
The uniqueness of insular biota increases its vulnerability to threatening processes
The inherent uniqueness of insular biota (e.g., endemic species, distinct traits, unique lineages) increases its overall vulnerability to global change drivers (Fernández-Palacios et al. 2021a). Islands are isolated, have complex topographies, are prone to extreme events (e.g., volcanic eruption, landslides, hurricanes), and harbour fewer potential refuges (compared to mainland landscapes), making them more exposed to threats (Russell and Kueffer 2019) (Figure 1A). Islands also host a large number of endemic species: up to 90% of endemic non-vagile taxa (i.e., non-flying vertebrates, seed plants, molluscs or arthropods) occur on islands (e.g., Madagascar, Antonelli et al., 2022). Endemics have small population sizes and restricted geographical ranges (sometimes limited to an archipelago, an island or even a single peak, volcano or cliff) (Fernández-Palacios et al. 2021b), increasing their extinction probability (Manes et al. 2021). An iconic example is the radiation of Hawaiian Honeycreepers, a group of small birds particularly vulnerable to invasive alien species and habitat modification (Box 1).
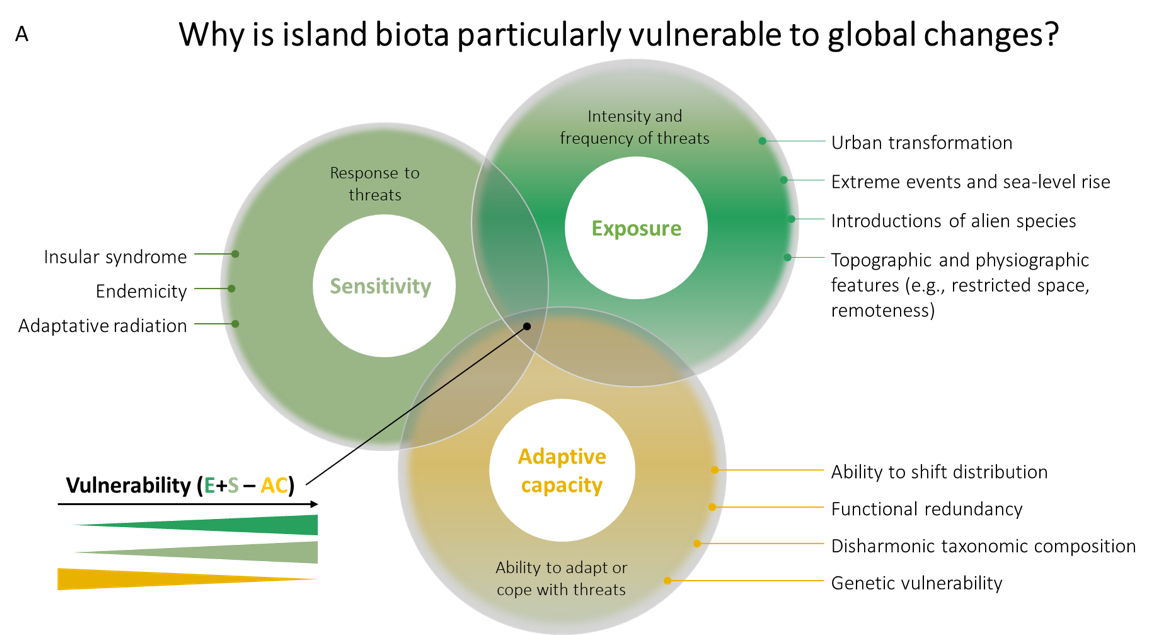
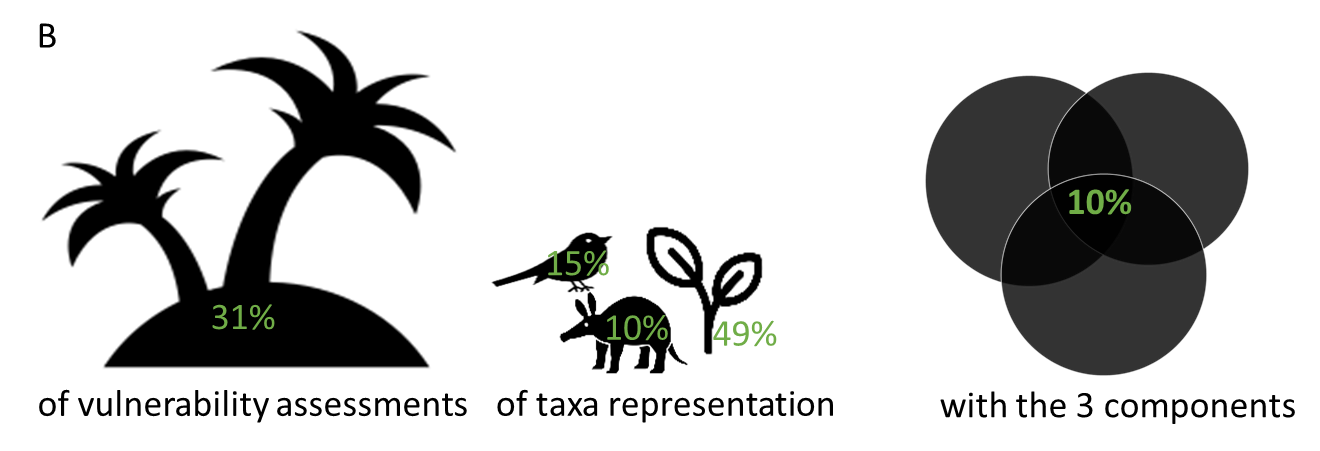
Figure 1 - A. Conceptual figure of the vulnerability of insular biota to global changes with a non-exhaustive list of the characteristics contributing to each vulnerability component (i.e., exposure, sensitivity, and adaptive capacity). For example, insular syndrome traits, endemicity, and adaptive radiation are characteristics that increase the sensitivity of insular biota to global changes compared to mainland biota, whereas (i) a lack of functional redundancy, (ii) disharmonic taxonomic composition, and (iii) genetic vulnerability are characteristics that decrease adaptive capacity. B. Percentage of island biota representation, taxonomic representation, and vulnerability components representation in current climate change vulnerability assessments. The numbers are calculated from de los Ríos et al. (2018).
Box 1 | The case of the Hawaiian honeycreepers
The Hawaiian Islands constitute an archipelago of eight larger and numerous smaller volcanic islands in the North Pacific Ocean, in total about 16,644 km2 in size, and about 3,200 km from the nearest mainland. The islands in the northwest are older and typically smaller, due to progressive erosion, while the islands in the southeast are larger and still volcanically active. Thanks to the archipelago’s extreme isolation and the islands’ variation in size and environmental conditions, Hawaii has been a hotspot of speciation and adaptive radiation (Baldwin & Sanderson, 1998; Lerner et al., 2011; Price, 2004). The Hawaiian honeycreepers (Fringillidae: Drepanidinae) represent one of the most iconic examples of adaptive radiation, which has resulted in a striking variation in bill morphology (Lerner et al., 2011). This strong adaptive radiation, however, has also led to species with highly restricted ranges and naturally small population sizes. Hence, little is needed to push these species over the brink of extinction and, to date, at least 36 of the 59 known species of honeycreepers are extinct (Figure 2a). Extinction has been non-random with respect to the functional trait space of the species, with extinct species generally larger in size (Figure 2b).
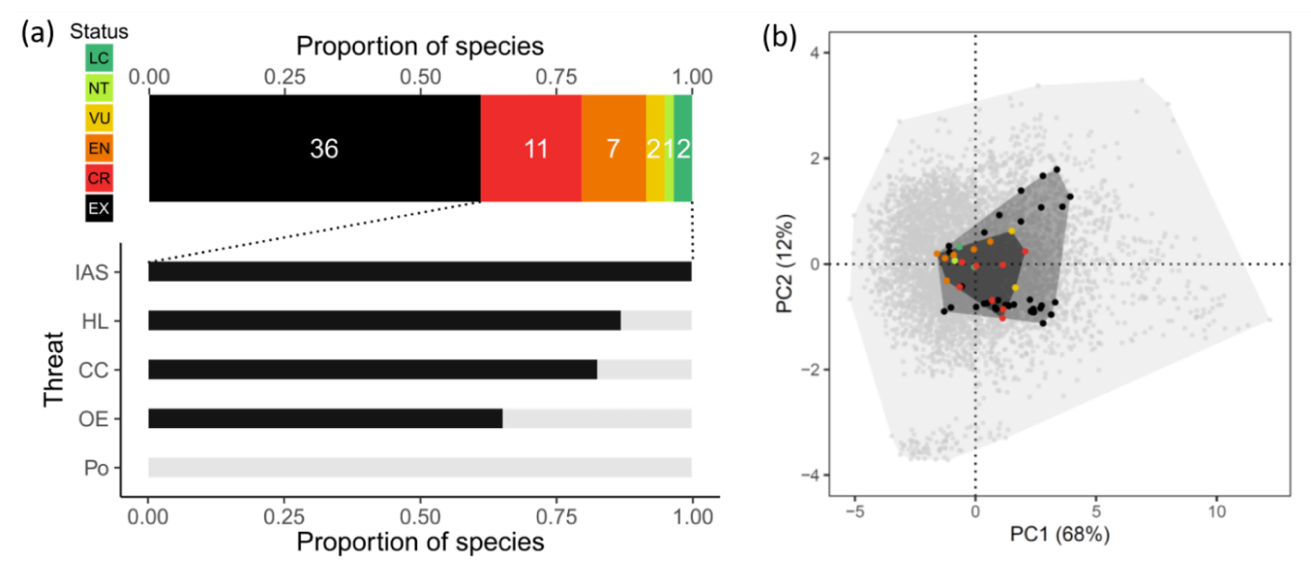
Figure 2 - Conservation status, threats, and morphospace of Hawaiian Honeycreepers. (a) IUCN Red List threat status of 59 known species of honeycreeper (LC: least concern, NT: near threatened, VU: vulnerable, EN: endangered, CR: critically endangered, EX: extinct) and prevalence of threats among the 23 extant species (IAS: invasive alien species, HL: habitat loss, CC: climate change and extreme events, OE: over-exploitation, Po: pollution) (b) The morphospace compared between 19 extant honeycreepers (dark grey polygon), 54 extant and extinct honeycreepers (medium grey polygon), and 5,974 extant passerine birds globally (light grey polygon). The morphospaces for the extant species were obtained through a principal component analysis (PCA) based on the eight morphological traits available in AVONET (Tobias et al., 2021), namely bill length from tip to skull along the culmen, bill length to nostrils, bill width and depth to nostrils, tarsal, wing and tail length, and Kipp’s distance. The same eight traits for the extinct honeycreepers were taken from (Matthews et al., 2023). The first two principal components explain 80% of the total morphological variance and are primarily associated with body size (PC1) and tarsus length versus Kipp’s distance (PC2), respectively. The list of species was obtained from (Matthews et al., 2023; Ricklefs, 2017) and the IUCN Red List of Threatened Species v2023-1. Species names were harmonized and duplicates removed based on the taxonomic backbone used by the IUCN.
The main drivers of past extinction, as well as current threats to the species, are the introduction of invasive alien species and the loss of habitat due to conversion to agricultural land. Invasive species include predators of birds and eggs (rats, cats, dogs), herbivores that modify the habitat (for example, the extinction of Laysan Honeycreeper Himatione fraithii is ascribed to the introduction of rabbits, which eliminated virtually all vegetation cover from the Laysan islands), and vector species of infectious diseases, such as avian malaria (Benning et al., 2002). Habitat loss began with Polynesian colonists, who cleared much of the low-elevation and seasonally dry forest for agricultural purposes, and was continued by later European colonists, who additionally converted high-elevation forests for pasturage (Riper & Scott, 2017). Climatic factors contributed little to past extinctions but are a progressive threat to extant species, especially because it may lead to an upslope shift of infectious diseases (notably avian malaria) (Benning et al., 2002). Overall, this example illustrates the importance of (i) considering multiple threats when focusing on island biota, (ii) taking into account the singularities of insular syndrome such as restricted ranges, small population size or level of endemicity, as well as (iii) the importance of considering markers of sensitivity at the community level given the non-random distribution of species at risk of extinction and their potential higher vulnerability to global change.
Insular species have also evolved distinct and unique ecological, physiological, behavioural, morphological, and life-history traits, a phenomenon commonly referred to as the ‘island syndrome’ (Baeckens and Van Damme 2020). These distinct and unique traits increase their sensitivity to current and future anthropogenic threats, including overharvesting, habitat loss, increased drought and invasive alien species (Zizka et al. 2022; Rozzi et al. 2023) (Figure 1A). This phenomenon is for example associated with dioecy in plants, which makes them more likely to disappear if their mutualists’ species go extinct, increasing their vulnerability to human-induced perturbations (Schrader et al. 2021). Reduced herbivory and predation pressure on islands leads to low levels of spinescence compared to their mainland counterparts, making island species more susceptible to the introduction of exotic herbivores (Burns, 2016; Clavero et al., 2009), unless they have coevolved with megafaunal herbivores (Barton et al. 2024). The loss of flight capacity renders insular birds unable to escape from anthropogenic threats (Sayol et al., 2021), making them more susceptible to extreme weather events, such as hurricanes, or gradual environmental change (e.g., progressive directional changes such as climate change) (Roff 1990; Burns 2019).
Additionally, the intricate and often rugged topography of oceanic islands frequently leads to long-term isolations of populations, which may result in genetic drift and then inbreeding when population sizes are small (Frankham, 2002). Furthermore, island colonization within archipelagos, and isolation by distance, could also lead to high population structure between islands (White and Searle 2007; Parmakelis et al. 2015). These populations are thus more prone to genetic diversity loss, which may result in fewer opportunities to adapt to changes. Although this does not necessarily increase their vulnerability per se, the loss of genetic diversity due to natural or anthropogenic disturbances, especially when exposed to disturbances over a short time period (Inamine et al. 2022), can cause a demographic or genetic collapse. Thus, losing an insular population can cause demographic and genetic loss, whereas it causes only demographic loss on the mainland. Because of that, the effective conservation units on islands should be, in most cases, at the population level, rather than species level (Melo & O’Ryan, 2007).
Besides population- and species-level characteristics, the composition of insular species’ assemblages harbours features that make them more vulnerable. One example is their disharmonic taxonomic composition, which refers to the systematic over- or under-representation of certain taxonomic groups compared to mainland assemblages. Notable examples include the absence of several families of non-volant mammals on many isolated islands (Brace et al. 2015), and the over-representation of pteridophytes in island flora (Kreft et al. 2010). The absence of certain functional groups on islands, although partially compensated by in-situ diversification and differentiation, allows incoming exotic species to fill vacant niches (Vitousek et al. 1997). These exotic species can exert new role (e.g., predation, competition) unknown to the native community, which is evolutionarily naive and functionally unequipped to withstand these novel pressures. Additionally, species traits in island communities are often complementary rather than redundant, leading to communities with low functional redundancy (Whittaker et al. 2014; Harter et al. 2015). This makes them more sensitive to threats (McConkey and Drake 2015) due to the lack of ‘ecological insurance’; the ability to replace the missing functions (Loreau et al. 2021).
State of the art of vulnerability assessments and their limitations when applied in insular contexts
Despite the inherent vulnerability of insular biota to global changes, most biodiversity vulnerability assessments have been designed for and focused on mainland systems. Climate change vulnerability assessments emerged in the 1990s to anticipate impacts and prepare appropriate responses (Foden et al. 2018). Most definitions concur that a species’ vulnerability to a threat is determined by three components: exposure, sensitivity, and adaptive capacity (Foden et al. 2013a; Foden et al. 2018; Butt et al. 2022). Using this definition, a recent review showed that among the 741 studies assessing climate change vulnerability, the majority focused on mainland systems, with less than one third (n = 231) including islands (de los Ríos et al. 2018) (see Table S2 for examples) and only 136 studies associated with a specific insular country. Although this would be representative of the small land surface area occupied by islands (6.7%), it falls short in terms of biodiversity representativeness, as island’s biodiversity represents 20% of biodiversity worldwide, with 50% of threatened and 75% of known extinctions (Fernández-Palacios et al. 2021b). These island vulnerability assessments are often geographically and taxonomically restricted towards high income countries (e.g., Australia, the UK, and the USA account for 60% of studies on insular biota) and plants (49% of studies) (Figure 1B), respectively. In addition, the large majority of vulnerability assessments do not consider the influence of multiple threats (but see Santos et al., 2021; Sousa et al., 2021; Ureta et al., 2022), which are likely to act together, interact with and be exacerbated by climate change (e.g., habitat loss, overexploitation, biological invasions), potentially leading to synergistic impacts (Leclerc et al. 2018). Note that these geographic, taxonomic, or conceptual biases occur in both mainland and insular assessments (see also de los Ríos et al., 2018). Finally, those assessments do not consider other dimensions of diversity, neither they take into account the specificities of insular biota (e.g., phylogenetic endemism, geographic isolation of islands, population size, etc.) that make them more likely to be vulnerable to global changes..
Assuming that exposure is a suitable proxy for the impact of a particular threat, most vulnerability assessments focused on exposure alone, with less than 10% considering the three components of vulnerability (Butt et al. 2016; de los Ríos et al. 2018). This assumes that all species will have equal responses to a threat, which is highly unlikely. In fact, the likelihood of species being impacted by certain threats, either on the mainland or on islands, is modulated by their traits (Leclerc et al. 2020a; Fromm and Meiri 2021; Marino et al. 2022; Soares et al. 2022). We argue that trait-based vulnerability assessments, when applied to a range of taxa and threats, can provide a useful approach for (i) developing a more comprehensive index of vulnerability to threats, and (ii) informing effective management actions for conservation (Gallagher et al. 2021).
An adapted framework for assessing the vulnerability of island biota to multiple threats
We present a conceptual framework divided into five steps that provides a roadmap for vulnerability assessments of biota in insular systems, considering the three components of vulnerability (i.e., exposure, sensitivity, and adaptive capacity; Figure 3). Here, we refer to islands as insular systems that have a landmass smaller than Greenland (i.e., < 2.17 million km²) and are surrounded by sea water. We built upon previous framework, which focused on taxonomic diversity in mainland ecosystems (Foden et al. 2013a; Parravicini et al. 2014; Leclerc et al. 2020b; Butt et al. 2022) to include in our framework various markers for each vulnerability component that are specifically tailored to the inherent characteristics of island biota, as well as multiple threats, taxa, and dimensions of diversity (i.e., taxonomic, functional and phylogenetic diversity). The development of this vulnerability framework tailored for island systems is part of a larger initiative to meet international policy targets that better integrate biodiversity threats and dimensions (Box 2).

Figure 3 - Conceptual framework for assessing the vulnerability of island biodiversity to global changes. The framework consists of five steps and can be iteratively applied (see circular arrow). Markers in bold font represent the markers specifically designed for insular biota but other markers are also important to indicate species’ sensitivity, independently of their occurrence on islands or mainland.
Step 1: Identify the scope and aim of the vulnerability of insular biota
The first step is to define the scope of the vulnerability assessment in terms of spatial and temporal extent, relevant threats, and studied biota (taxonomic group, biological level, biogeographical origin of species). This challenging task is pivotal for the assessment design, which in turn affects the final step of informing conservation actions (Step 5). For example, broad-scale assessments (e.g., at the global extent or among several archipelagos) contribute to strategic planning and to establish a common baseline of vulnerability information (e.g., IPBES assessments), while local-scale assessments (e.g., within archipelagos or group of islands) are appropriate for informing site-level management decisions.
For instance, a vulnerability assessment could be conducted at the spatial extent of a national park, within an island of a few hectares only, with a restricted set of species (e.g., Harper et al., (2022), 24 ha in South Africa, 18 amphibian and 41 reptile species). This can inform management priorities at the landscape scale, such as defining park-use zones to help allocate restricted areas acting as corridors for species migration, or creating habitat conditions for breeding (Harper et al., 2022). At this level, an explicit treatment of population genetics and/or population viability analyses could also be conducted, and this may become more feasible in the future with the emergence of macrogenetics studies (Leigh et al. 2021). In parallel, studies focusing on the global extent are key to assessing vulnerability metrics, identifying geographical shortfalls in data coverage, and supporting the implementation of conservation policies to mitigate biodiversity losses. Note that, in all cases, controlling for island area or species richness is essential to mitigate biases towards larger islands when calculating vulnerability metrics. Finally, even if vulnerability assessments are conducted at the global scale, particular attention should be directed toward endemic species or insular populations, since island biodiversity conservation operates at the population level.
Step 2: Determine the markers of exposure, sensitivity, and adaptive capacity
There are many different relevant markers associated with biodiversity’ vulnerability to threats. Those markers may vary in type (e.g., ecological or demographic) and organisation level (e.g., population, species, community, or ecosystem), and could be considered either simultaneously or separately in the framework. Literature review and expert opinion can be used to identify and collect relevant markers for each component of vulnerability based on the scope and purpose of the assessment. For instance, assessing the vulnerability of terrestrial species to land-use change and climate change requires markers of exposure to climate change (e.g., change in precipitation regimes, sea-level rise) and land-use change (e.g., urbanization rates, forest conversion into agriculture, infrastructure development, shift from agriculture to tourism) (e.g., Bellard, et al., 2015a; Bellard, et al., 2015b). Moreover, it is necessary to list the biological traits related to the sensitivity and adaptive capacity of the studied insular species to the above-listed threats (Figure 3 and Table S1 for examples of exposure, sensitivity, and adaptive capacity markers). Markers of exposure can be linked to the focal threat(s) or general global changes, and multiple markers should be used to capture the multiple dimensions of changes (e.g., temperature change, heat waves, droughts).
Species’ sensitivities are determined by their intrinsic traits (e.g., climatic niche breadth, habitat specialization, diet breadth, body size, etc.). Despite the increasing availability of traits for birds, mammals, reptiles, and plants (Faurby et al. 2018; Tobias et al. 2021; Soria et al. 2021; Díaz et al. 2022), trait-based approaches have been barely applied to date or they are specific to some threats. Some sensitivity traits might be specific to a given threat (e.g., temperature tolerance), while others are general and encompass sensitivity to different threats (e.g., body size, dependence on interspecific interactions). Note that for both sensitivity and adaptive capacity, a comprehensive list of markers is applicable at either the population or species level (see Thurman et al., 2020 for a list and Table S1). However, markers applicable at the community level, such as functional or phylogenetic diversity, could also be included (Table 1 for example). Using extinctions scenarios for threatened species, we can also establish which islands are more likely to lose a significant share of their functional and phylogenetic diversity (richness, redundancy, originality, uniqueness) (Bellard et al. 2021; Llorente-Culebras et al. 2024). Those analyses could help identify islands that are more likely to be sensitive at the community level.
Adaptive capacity refers to the ability of insular biota to respond to stressors, caused by multiple threats or novel conditions, by either persisting in situ or by shifting in space or time (following Thurman et al., 2020). Acclimation, behavioural change, phenotypic plasticity and evolutionary adaptation may all contribute to adaptive capacity (Foden et al. 2018; Royer-Tardif et al. 2021). For mobile animals, especially the most vagile organisms, markers of adaptive capacity can incorporate traits linked to movement or mobility (e.g., migration frequency and distance, site fidelity) (Thurman et al. 2020; Butt et al. 2022). Note that for plants and sessile organisms in general, adaptive capacity mostly refers to their capacity to persist in situ with mating system and fecundity as strong markers (Thurman et al. 2020). Finally, adaptive capacity of insular biota can also be modulated by extrinsic factors, such as habitat quality, habitat availability, habitat connectivity and level of protection, factors which could therefore be included in the measure of adaptive capacity at the system level (i.e., island, archipelago).
Table 1: Proposed functional-based and phylogeny-based metrics that could be used as markers at the community level in vulnerability assessments.
|
|
Functional-based markers |
Phylogeny-based markers |
|---|---|---|
|
Sensitivity |
Functional rarity |
Phylogenetic endemism |
|
Definition - Functional rarity, expressed at the species level, is the combination of the functional distinctiveness/uniqueness (based on traits), the scarcity and the geographic restrictedness of a species (Violle et al., 2017). |
Definition - Phylogenetic endemism is the spatial restriction of the phylogenetic diversity of a community (Rosauer et al., 2009). This is a relative measure of endemism that represents the degree to which lineages or branches of the tree of life are spatially restricted. |
|
|
Rationale - Given that functionally rare species can play a critical role in ecosystem functioning, and because rarity is linked to species’ sensitivity (Davies et al., 2004; Loiseau et al., 2020), areas with a high proportion of functionally rare species are particularly susceptible to threats. |
Rationale - Areas with high phylogenetic endemism are characterized by the presence of species that have diversified and evolved in response to specific environmental conditions within a particular location. Among the drivers shaping phylogenetic endemism, climate plays a significant role (Guo et al., 2023), implying that changes in climate conditions may disproportionately affect these species, potentially leading to their decline or extinction. |
|
|
Adaptive capacity |
Functional redundancy |
Phylogenetic distinctiveness |
|
Definition - Functional redundancy of a given community reflects the tendency for the constituent species to perform similar functions (Mouillot et al., 2014). |
Definition - Phylogenetic distinctiveness reflects the degree of isolation of a species or a group of species within a phylogenetic tree (Pavoine & Ricotta, 2021). |
|
|
Rationale - Functional redundancy has been theoretically and empirically linked to the concepts of resistance and resilience of ecosystem functioning to species loss (Biggs et al., 2020) via the insurance hypothesis (McCann, 2000). Hence, more functionally redundant systems should show greater resilience to perturbations (Mouillot et al., 2014) become role of extinction species can be fulfilled by functionally close species. This indirectly reflects the ability of the system to adapt to disturbances. |
Rationale - Evolutionarily distinct species or group of species represent uniquely divergent genomes (Warren et al., 2008). Consequently, sets of evolutionarily distinct species are expected to encompass a large proportion of the parental clade’s total phylogenetic diversity, which may play a crucial role in ensuring long-term stability and resilience (Cadotte et al., 2012). This implies that communities exhibiting higher phylogenetic distinctiveness are more likely to harbor increased evolutionary potential, enabling them to adapt. |
Step 3: Compute measures of vulnerability and its components
Once all the markers of the three components of vulnerability are collected, the next step is to combine them all into a composite vulnerability measure. However, aggregating multiple markers into a single value for each of the three vulnerability components is challenging. A recent review showed that most assessments use arithmetic mean for the aggregation (Tonmoy et al., 2014). In the case of quantitative markers, as here, one possibility is to normalise each marker to a 0-1 range. This transformation, which can be done with multiple methods (e.g. Leclerc et al., 2020a), creates unitless metrics with equal weight. Then the markers of each component (i.e., exposure, sensitivity, and adaptive capacity) can be summed and re-scaled to obtain normalized values of exposure, sensitivity, and adaptive capacity. This technique has the advantage of clearly identifying which components drive vulnerability, by ensuring that all markers are weighted equally, and can thus effectively guide the implementation of conservation actions at the island level. The different markers could also be weighted differently to put more emphasis on specific markers depending on the current level of island protection policy or biodiversity richness occurring on each island. Finally, when all the data have been aggregated, the majority of the current methods employ criteria-based approaches, classifying biota into categories of vulnerability from low to high by summing up the different components of vulnerability (hereby referred to as qualitative or semi-quantitative frameworks).
To avoid arbitrary thresholds, we propose a quantitative framework using multicriteria decision analysis (Leclerc, et al., 2020a; Parravicini et al., 2014) that provides continuous vulnerability values and a relative ranking of islands or archipelagos. This method ranks alternatives according to their distance to positive (i.e., minimal vulnerability, with low exposure and sensitivity, and high adaptive capacity) and negative ideal solutions (see Leclerc et al. (2020b) for details). Note that alternative approaches exist for combining the three vulnerability components, such as additive effect between components (Nyboer et al. 2021) or interacting effect between exposure and sensitivity for instance (Silva Rocha et al. 2024). In any case, we recommend carrying out robustness analyses to explore the uncertainty potentially introduced by the aggregation and weighting methods.
Step 4: From uncertainty assessments to the improvement of vulnerability assessments and policy recommendations
The outcomes of vulnerability assessments can be affected by several factors, including missing data, variation of underlying traits, aggregation techniques, and, in the case of future-looking assessments, by uncertainties about the future trajectories of anthropogenic threats. Techniques to estimate uncertainty from missing trait data (e.g., Hossain et al., 2019) as well as uncertainty in modelling the future of biodiversity (e.g., Thuiller et al., 2019) are readily available. In addition, we recommended carrying out robustness analyses to assess the impact of including or excluding specific markers from the calculation, by rerunning the vulnerability assessment without the focal marker. Robustness analyses may also be used to explore the effects of alternative aggregation or standardisation techniques (Tonmoy et al., 2014; see also Boyce et al., 2022). We also recommend validating the outcome of the vulnerability assessments with data from another temporal period (e.g., by comparing with past data), or by comparing with other islands using documented vulnerability or other metrics of vulnerability (e.g., IUCN Red List of species, Living Planet Index). In this context, it is crucial to both estimate and communicate the uncertainty. This can be achieved by, for example, mapping the variance of the estimated vulnerability metric or using maps of ignorance (e.g., Rocchini et al., 2011; Tessarolo et al., 2021). As well as identifying future research priorities, uncertainty evaluations strengthen confidence in assessments, thereby increasing their uptake to support policy decisions, including by the IPCC and IPBES. We also advocate for the integration of interactive interfaces and dashboards with options to select threats, markers, taxa, or ecosystems of interest, as well as different narratives trajectories, which may complement the scientific message - particularly helpful for simplifying multidimensional information (McInerny et al. 2014). Finally, we recommend conducting an ongoing review and iteration to enable adaptation and improvement of the vulnerability framework including as new ecological data become available (Henry et al. 2024).
Step 5: From vulnerability assessments to conservation actions and policy
Effective conservation practice under global change relies on identifying which species are most vulnerable and what drives their vulnerability (Box 2, Step 5). Vulnerability assessments such as those described in Steps 1-4 provide invaluable information for conservation priority-setting for island biodiversity. For species of the highest vulnerability, for example, the specific traits and exposure measures driving the vulnerability can inform appropriate responses (e.g., captive breeding, assisted translocation, disease management). For species identified as sensitive to climate change, but which may escape impact through high adaptive capacity, such responses (e.g. range shifts, phenological change) should be facilitated and monitored (Foden et al 2013a). Species with poor adaptive capacity but low sensitivity can be assumed to be able to withstand change in situ, but close monitoring should be maintained to ensure that this assumption is realised (Foden et al, 2013a) and that unexpected thresholds are not crossed. Assessments can also inform landscape-level actions (e.g. protecting hotspots of high numbers of vulnerable species, habitat restoration, and increasing permeability) (Harper et al. 2022).
Biodiversity policy too may be guided by species vulnerability assessments. For instance, one of the targets of the KM-GBF is to minimize the impact of climate change on biodiversity through (among others) mitigation, adaptation and risk disaster reduction actions. Such aims require the use of different markers of adaptive capacity through multiple plausible scenarios (e.g., protected areas, habitat intactness) to inform how different management actions can impact the vulnerability. Vulnerability assessments can help target the control and eradication of invasive alien species in conservation priority sites such as islands (Target 6), and also identify the sites that need to be brought under protection (Target 3) to conserve biodiversity. Monitoring progress in meeting these conservation targets too may be informed by vulnerability assessments. Since species sensitivity and adaptive capacity traits change little over time, while exposure increases at an observable rate, changes in vulnerability may also provide a measure of the relative effectiveness of conservation actions implemented (Foden et al. 2013b). Because the exposure component is common across species (e.g., inundation, invasive alien species), the threat itself may be managed through dedicated programs, such as mangrove rehabilitation or invasive alien species control (Jones et al. 2016).
Finally, beyond its use in current policies, our vulnerability framework could be useful in drafting new policies. Indeed, to set relevant targets in the new policies being drafted (e.g., by [year], reduce by [number] % the number of species threatened by [threat]), policymakers need to know (i) the current situation and (ii) what could be the future situation under different scenarios. The proposed vulnerability framework has the potential to provide both aspects and may contribute to the establishment of policy targets that are ambitious and attainable for insular biodiversity.
Box 2 | Leveraging an insular vulnerability framework to inform multiple policies
Our proposed vulnerability framework for insular biodiversity is particularly relevant for informing various vulnerability policies (in orange) and biodiversity policies (in green) at both international and supranational levels, such as the European Union (EU).
First, by quantifying the three components of island vulnerability, our framework can directly provide the currently missing biodiversity component of the Multidimensional Vulnerability Index (MVI). The MVI is developed by the Small Island Developing States (SIDS; https://www.un.org/ohrlls/mvi/) with the United Nations to characterise the financing help for sustainable development needed in the face of global changes (arrow 1, Figure 4). The SIDS is a political coalition of 39 low-lying islands that are united by the threat posed by climate change to their survival. In climate change negotiations, they are a loud and powerful voice for upscaling climate action, since they are disproportionally affected by climate change consequences. To understand and respond, several assessments of these islands’ climate change vulnerability have been carried out (e.g., UN High Level Panel, 2022; UNFCCC, 2007), but none includes more than a broad mention of biodiversity impacts. A scientifically rigorous assessment of climate change vulnerability of island biodiversity is, therefore, both extremely important and critically urgent. Our framework can also help EU Member States, which are required to report to the European Commission on their disaster risk management activities, to identify which components of vulnerability they can act on (arrow 2). The risk of biodiversity loss has been recently included in the Recommendations for National Risk Assessment for Disaster Risk Management in the EU (Poljansek et al., 2021), which does not specifically cover insular biodiversity. In that context, our framework can provide real added? value for EU Member States with insular territories. Our framework can also further help identify whether mitigation strategies for islands to reach Target 8 of the Kunming-Montreal Global Biodiversity Framework (hereafter, GBF) should focus on limiting territories’ exposure or promoting the adaptive capacity of insular biota (arrow 3).
Second, our framework can help EU Member States with insular territories to identify and prioritise which species, sites and threats they should use to best mitigate their vulnerability in the context of their National Risk Assessment (arrow 4). Such actions can also contribute to (i) identifying the species, sites and to focus the management measures to reach Target 4 of the GBF (arrow 5), (ii) determining the most vulnerable islands to invasive alien species to reach Target 6 of the GBF (arrow 6), and helping EU Member States with insular territories to identify species threatened by invasive alien species on which to focus conservation actions to reach Target 12 of the EU Biodiversity Strategy for 2030 (https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A52020DC0380 EU BDS, (arrow 7)).
Finally, by providing multiple scenarios of threat trajectories and a list of priority islands for conservation action, our framework can contribute to documenting the risk of biodiversity loss under different conservation scenarios to EU Member States with insular territories (arrow 8). The vulnerability framework could contribute to identifying priority islands for restoration (target 2 of the GBF and target 4 of the EU BDS (arrow 9)) and for protection (target 3 of the GBF and target 1 of the EU BDS (arrow 10)), as well as in determining which invasive alien species should be eradicated or controlled (target 6 of the GBF (arrow 11)).
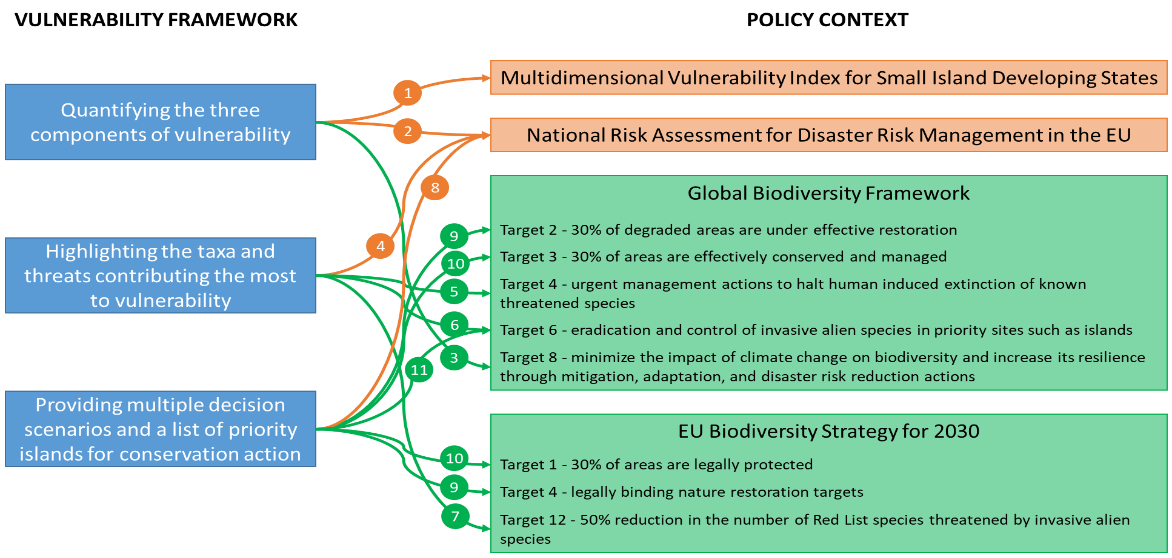
Figure 4 - This figure highlights some possible outcomes of the vulnerability framework and how they are linked (arrows) to the vulnerability (in orange) and biodiversity policies (in green). Each circled number represents a concrete example of how our vulnerability framework can support existing policies, and all those examples are detailed in the box.
Concluding remarks
Given the proliferation of multiple threats that islands are facing, efforts must be made to study island biota in the light of global changes, and thus to embrace the whole of biodiversity, mainland and insular, in current vulnerability assessments in international arenas such as the IPCC or IPBES. Our comprehensive and detailed framework lays the foundations for understanding and predicting island biodiversity vulnerability to global change. Besides, this framework aims to integrate island-specific characteristics with enhancements to existing frameworks that may also prove beneficial for continental assessments. These enhancements include the consideration of multiple threats, indicators that reflect phylogenetic and functional diversities, and an uncertainty analysis that is essential for all types of vulnerability assessments. Because the outcomes of vulnerability assessments will be challenged by missing data and, in the case of future assessments, by uncertainties about the future trajectories of anthropogenic threats, it is crucial to both estimate and communicate uncertainty (e.g., Hossain et al., 2019; Rocchini et al., 2011; Tessarolo et al., 2021; Thuiller et al., 2019), which will ultimately help to protect biodiversity with more robust information.
Glossary
Adaptive capacity: the ability of a population or species or community to adapt to changing conditions; this may be via ecological (i.e., physiological and/or behavioural plasticity) or evolutionary adaptation (i.e., through natural selection acting on traits).
Adaptive radiation: the rapid diversification of a single evolutionary lineage into multiple ecologically or morphologically distinct species.
Exposure: the extent to which each population or species’ or a community physical environment changes due to global threats (Foden et al., 2013a). It includes the intensity, magnitude and frequency of the threat(s).
Sensitivity: the intrinsic capacity of population or species or community to cope with threats, based on their life-history, ecology, morphology, or behaviour.
Threat: external factor that has the potential to impact the viability, abundance, distribution, or behaviour of a population, species, or community.
Vulnerability: susceptibility of a system/species to a negative impact following exposure to a threat
Acknowledgements
We are grateful to all the participants of the project and workshops (scientists and support staff of the CESAB). Preprint version 4 of this article has been peer-reviewed and recommended by Peer Community In Ecology (https://doi.org/10.24072/pci.ecology.100697; Dakos, 2025)
Data, scripts, code, and supplementary information availability
Data and scripts are available online at https://doi.org/10.5281/zenodo.15350333 (Cbellard, 2025).
Supplementary information is available online at https://doi.org/10.5281/zenodo.15350239 (Bellard, 2025).
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Funding
This publication is a product of the RIVAGE project
co-funded by the synthesis center CESAB of the French Foundation for Research on Biodiversity (FRB; https://www.fondationbiodiversite.fr).