
 CC-BY 4.0
CC-BY 4.0
Introduction
Human modified environments are rapidly increasing (Goldewijk, 2001; Liu et al., 2020; Wu et al., 2011), which puts other species in the precarious position of either adapting to the new challenges or moving to a different area if they are not able to adapt (Sol et al., 2014, 2017). Behavioral flexibility (hereafter, ‘flexibility’), the ability to change behavior when circumstances change via processing information that becomes available to other cognitive operations (see Mikhalevich et al., 2017 for theoretical background), is hypothesized to play an important role in the ability of a species to adjust to new areas and rapidly expand its geographic range (Chow et al., 2016; Griffin & Guez, 2014; Lefebvre et al., 1997; Sol et al., 2002; Wright et al., 2010). The prediction is that flexibility, along with behaviors such as exploration, and innovation, facilitate the expansion of individuals into completely new areas, and that the role of these behaviors diminishes after an initial adjustment stage (Wright et al., 2010). This prediction is supported by experimental studies showing that abilities that are not commonly (or ever) observed in the wild are primarily expressed when needed (Auersperg et al., 2012; Bird & Emery, 2009; Laumer et al., 2018; Manrique & Call, 2011; e.g., Taylor et al., 2007). Therefore, the founding individuals who initially dispersed out of their original range should not have unique behavioral characteristics that are passed on to their offspring. Instead, the continuation of a range expansion should rely on flexibility, and potentially behaviors such as exploration, innovation, and persistence. These behaviors should therefore be expressed more on the edge of the expansion range where there have not been many generations to accumulate relevant knowledge about the environment, and expressed more in species and populations that are rapidly expanding their range relative to species that are not.
Whether and how flexibility relates to a rapid range expansion is still an open question (Wright et al., 2010). To determine whether behaviors are involved, they must be directly measured in individuals in populations across the range of the species, and, ideally, also include cross-species comparisons using the same methods (see a discussion in Logan et al., 2018 and direct evidence in Logan et al., 2025 on the danger of flexibility proxies). There is only a small amount of direct evidence that is beginning to answer this question and it suggests that populations on the range edge express certain behaviors more than populations away from the edge. Magory Cohen et al. (2020) showed that common mynas (Acridotheres tristis) in populations on the edge were more innovative and less neophobic with food than individuals in populations away from the edge, while there were no differences in object neophobia. Logan et al. (2023b) found that edge great-tailed grackles (Quiscalus mexicanus; hereafter GTGR) were more persistent and had a higher flexibility variance than individuals in a more central population, while there were no differences in average flexibility, innovation, or exploration. Evidence in invasive species in regions where they were introduced also shows that individuals on the edge or in newer populations were faster to eat new foods (house sparrows, Passer domesticus; Martin & Fitzgerald, 2005; Liebl & Martin, 2014) and were more risk averse (bank voles Myodes glareolus; Eccard et al., 2023) than individuals away from the edge or in older populations. In contrast, invasive spiders (Cyrtophora citricola) at newer sites in their non-native range were less exploratory and less bold than spiders at older sites (Chuang & Riechert, 2021). Such differences can also vary seasonally: Liebl & Martin (2012) found that invasive house sparrows closer to the edge of their non-native range were more exploratory than those away from the edge, but only during the breeding season. This suggests that behavior is differentially involved in expanding a range, however flexibility may not play the primary role and the relative expression of the behaviors might be contrary to predictions. Most evidence on this topic comes from invasive species in their non-native range. GTGR are not technically considered invasive because their rapid range expansion is not due to human introductions (see Logan et al., 2023b for discussion). However, it is useful to compare GTGR with invasive species because the range expansion dynamics after arrival appear similar (Chapple et al., 2012).
While some intra-species comparisons of behaviors in edge and non-edge populations exist, we were not able to find inter-species investigations that directly measure flexibility or similar behaviors in closely related species that differ in their range expansion rates. This comparative method is a useful approach because it can serve as a type of natural experiment when testing closely related species that are known to differ in one of the two traits of interest (Davies et al., 2012). Here, we investigate flexibility in two closely related species, GTGR and boat-tailed grackles (Quiscalus major, hereafter BTGR), that differ in their range expansion rates: GTGR are rapidly expanding, while BTGR are not (DaCosta et al., 2008; Wehtje, 2003). GTGR are highly flexible (Logan, 2016; Logan et al., 2023a), but no flexibility data exist for BTGR. Both species are associated with human-modified environments, and are social and polygynous. Both species eat the same kinds of foods: a variety of human foods in addition to foraging on insects and on the ground for other natural food items (Johnson & Peer, 2001; Post, 1992; Post et al., 2020).
We previously found that GTGR individuals that were faster at reversal learning, a common method to quantify flexibility, had more food type switches during their focal follows (Logan et al., 2023b) and were faster at switching between options on a puzzlebox (Logan et al., 2023a). Using food type switches as a measure of flexibility is theoretically similar to using switches between options on puzzleboxes as a measure of flexibility in that individuals need to consider their options and decide when to switch and what to switch to (e.g., Logan et al., 2023b). Food type switching also more directly reflects whether individuals rely on flexibility when dealing with the challenges in their environment (Wiggins et al., 2006). We aimed to compare flexibility, measured as food type switches during focal follows, in two populations of GTGR (an older population where they have been breeding since 1951 (Wehtje, 2003) in the middle of the northern expansion front, Tempe, Arizona, and a recent population where they have been breeding since 2004 (Pandolfino et al., 2009; Yolo Audubon Society’s newsletter The Burrowing Owl July 2004) on the northern edge of the expansion front: Woodland, California) with one population of BTGR in the center of their range (Venus, Florida; the age of the population is unknown, but likely thousands of years old; DaCosta et al., 2008). The comparison of flexibility across these three populations first allows us to confirm that flexibility is generally high across the range of GTGR, which is what we previously found using reversal learning, and second to determine whether BTGR have lower flexibility than GTGR in general or GTGR at the edge (GTGR-GTGR population comparison predictions are in Logan et al., 2023b).
Determining whether GTGR are more flexible, particularly on the range edge, will allow us to accumulate more evidence on whether flexibility might play a key role in the GTGR rapid geographic range expansion. Alternatively, if BTGR and GTGR perform similarly, this would suggest that other abilities, ecological, behavioral, or physiological, may play a larger role than flexibility in restricting the BTGR range expansion.
Preregistered research question
Are there differences in behavioral traits (flexibility, innovation, exploration, and persistence) between BTGR and GTGR? (See Table 1).
Prediction: If behavior modifications are needed to adapt to new locations, then GTGR, which are rapidly expanding their geographic range (Wehtje, 2003), will have higher averages and/or larger variances than BTGR, which are not rapidly expanding their range (Wehtje, 2003), in at least some behavioral traits (e.g., behavioral flexibility: speed at reversing a previously learned color preference, innovativeness: number of options solved on a puzzle box, exploration: latency to approach/touch a novel object, and persistence: proportion of trials participated in). Higher averages in behavioral traits indicate that each individual can exhibit more of that trait. If resources are regularly distributed in time and space, perhaps BTGR require less flexibility when visiting these resources and attend less to their temporal availability or the individual’s food preferences. Perhaps the problems BTGR solve do not require much exploration or persistence. Lower variances in behavioral traits indicate that there is less diversity of individuals in the population, which means that there is a lower chance that some individuals in the population would innovate new foraging techniques and be more flexible, exploratory, and persistent if the population average is low.
Prediction alternative 1: Human-modified environments are suitable habitat for GTGR and BTGR and the amount of human-modified environments has been increasing. If the original behaviors exhibited by these species happen to be suited to the uniformity of human-modified landscapes, then averages and/or variances of these traits will be similar in the GTGR and BTGR sampled. This supports the hypothesis that, because these species are closely associated with human-modified environments (Wehtje, 2003), which may be similar across the geographic range, individuals in new areas may not need to learn very much about their new environment: they can eat familiar foods and access these foods in similar ways across their range (e.g., fast food restaurant chains likely make the same food and package it in the same packaging in Central and North America, outdoor cafes and garbage cans also look the same across their range).
Prediction alternative 2: If BTGR have higher averages and/or larger variances in the behavioral traits measured, this could indicate that perhaps these traits are not the primary facilitators of the GTGR’s rapid geographic range expansion. Alternatively, perhaps these species differ in a life history variable that restricts the BTGR from expanding, or there is some geographic feature that prevents the BTGR from rapidly expanding its range.
Study design table
Table 1 - The Stage 1 did not have a study design table because it was written before PCI RR existed and was only transferred to PCI RR in 2025. Therefore, we include a study design table for Stage 2.
Question | Hypothesis | Sampling plan | Analysis plan | Rationale for deciding the sensitivity of the test for confirming or discofirming the hypothesis | Interpretation given diferent outcomes | Theory that could be shown wrong by the outcomes | Result |
Are there differences in flexibility between boat-tailed and great-tailed grackles? | Food type switches average and variance: GTGR > BTGR | Bespoke Bayesian analysis in Logan et al. 2023b showed that we were able to detect differences between populations with sample sizes of 6 and 7. We are using this exact analyses in the current study | Bayesian model: Response: Number of food type switches per total number of seconds of observation time for each bird Explanatory: Population (BTGR, GTGR Arizona, GTGR California) (See Analyses section for more details) | Contrasts will determine whether the populations differ from each other. We will conclude that there is a difference if 89% of the difference between two sites is on the same side of zero | GTGR are more flexible than BTGR, and this could be a main facilitator of their range expansion | Non-behavioral traits are primary facilitators of rapid range expansions (Summers et al. 2023) | |
Food type switches average and variance: GTGR = BTGR | The original flexibility exhibited by these species happens to be suited to the uniformity of human-modified landscapes | Flexibility facilitates adapting to environmental change (see references in introduction) | There were no population or species differences in food type switches | ||||
Food type switches average and variance: GTGR < BTGR | Flexibility is not the primary facilitator of the GTGR range expansion | Flexibility facilitates adapting to environmental change (see references in introduction) |
Material and methods
Updates and changes to the preregistration
This study began as a preregistration, which received in principle acceptance at PCI Ecology in 2021: https://shorturl.at/otxtI. The preregistration contains the pre-planned analyses. Here, we report the rationale for the ways in which we conducted the study differently from the plan, and then describe the methods we used to obtain the results.
Changes made in the middle of data collection
After the first BTGR field season in 2022, we stopped the aviary experiments and switched to measuring reversal learning in the wild. We received permission to make the change in Feb 2023 from the PCI Ecology Managing Board and from the recommender, Esther Sebastián González, who incorporated it into our in principle acceptance. The reason for stopping the aviary experiments is that we discovered that BTGR are less robust to captivity compared to GTGR. BTGR seem to be having health issues as indicated by multiple deaths in the wild (including two that we witnessed), which unfortunately also carried over to the aviaries where two of the six aviary birds died. This meant that we were not able to obtain measures for three of the four variables we planned on investigating: innovation, exploration, and persistence. These three variables would have required aviary testing to conduct a comparison between the two species because this is the setting in which the GTGR were tested. We planned to measure reversal learning in the wild using an automated feeder experiment in GTGR (as in Logan et al., 2022). After data collection started in November 2023, and after extensive work trying to get the automated feeders to function, we discovered in January 2024 that the feeders were not usable. Therefore, we have no data on reversal learning in BTGR.
We previously measured flexibility in the wild in GTGR using behavioral observations (Logan et al., 2025), and found a variable that correlates with reversal learning performance: switching between food types during focal follows. This provided us with a way to collect comparable data in BTGR in the wild. Therefore, we added this measure of flexibility. Before we started collecting this data using focal follows, we received permission to make the addition in September 2023 from the recommender, Esther Sebastián González, who incorporated it into our in principle acceptance.
Sample
GTGR were caught in the wild in Woodland and Sacramento, California, and Tempe, Arizona; and BTGR were caught in the wild in Venus and Lake Placid, Florida. We aimed to bring adult grackles, rather than juveniles, temporarily into the aviaries for behavioral choice tests to avoid the potential confound of variation in cognitive development due to age, as well as potential variation in fine motor-skill development (e.g., holding/grasping objects; early-life experience plays a role in the development of both of these behaviors; e.g., Collias & Collias, 1964, Rutz et al. 2016) with variation in our target variables of interest. After switching away from the aviary tests in BTGR (see above), we continued to focus our study on adult individuals, however, it was not usually possible to discern female adults from juveniles. Adult GTGR were identified from their eye color, which changes from brown to yellow upon reaching adulthood (Johnson and Peer 2001). Juvenile male BTGR were identified by their dark brown feathers (rather than shiny black as in the adult males), and we were not able to distinguish between adult and juvenile females because they both have light brown feathers and brown eyes. We applied colored leg bands in unique combinations for individual identification. For some BTGR individuals, one leg band contained an RFID/PIT tag (Eccel Technology Ltd, https://eccel.co.uk/). GTGR were trapped in the wild using mist nets, walk-in traps, and bownets, and BTGR were trapped using walk-in traps. The bird was then processed by collecting biometric measurements, and, in the case of GTGR, also feathers and blood. After processing, the bird was released back to the wild either immediately (most BTGR and many GTGR), or after participating in behavioral tests in aviaries (some BTGR and many GTGR; the GTGR data are reported in other publications).
We first collected data on the GTGR populations (Logan et al., 2025), where we had a large sample because we were collecting data for many other purposes as well, including focal observations after individuals were released back to the wild. Whereas, for the BTGR, we were collecting data for this one purpose of comparing food type switch rates. Therefore, we set the minimum sample size (n=6) for the BTGR to a known sample size in the GTGR in which we were able to distinguish differences between populations.
The dataset consisted of eight BTGR and 76 GTGR (54 in Arizona, 22 in California) who had data that involved eating at least one food type. We met our minimum sample size of at least six individuals in each population and can therefore assess whether a given population is different from the others. A total of 36 food types across all sites were documented. The BTGR took a total of 14 food types, including cheese and oil, which the GTGR did not have. The Arizona GTGR took 20 food types, including lizard, bird poop, candy, vomit, condiment, and carcass, which the other populations did not have. The California GTGR took 15 food types, including mulch, which the other populations did not have.
Focal follow protocol
To calculate the probability of switching between food types in BTGR, we used the GTGR foraging behavior focal follow protocol from Logan et al. (2025; see Altmann, 1974 for a general description of the focal follow method). Ten minute focal follows were conducted on individuals in the wild between 9:00 and 16:00 in which all food types were recorded (both species), as well as the foraging bout start and end times (BTGR; Table 2). BTGR focals could only begin if the individual was in the foraging state (whereas, GTGR focals could begin at any time because we collected non-foraging data as well). This meant that we maximized our BTGR focal time by ensuring foraging behavior would be present in the focal (rather than starting a focal when they were sitting in a tree and maybe would not forage at all for the next couple of hours). We were careful to keep a respectable distance of at least 10m unless the bird approached us (using flight initiation distances in Møller, 2008 and Eason et al., 2006 as a baseline). If the focal individual showed alarm behaviors due to the observer (e.g., alarm calling at the observer), the focal was stopped and began again on a different day. We determined which individual to follow next by using the order in which the birds were caught. Of the color marked individuals at a given location that were currently in a foraging state, we conducted the focal on the one who was next on the list. If some individuals already had focal follows, we prioritized following those individuals next on the list who had fewer focal follows. If a focal follow was conducted in a particular location on a given day, no further focals could be conducted at that location on that day to avoid pseudoreplication of foraging behavior among individuals. The BTGR field site consisted of two distinct areas: one at Lake June Park in Lake Placid, Florida, a large 16 hectare park that we divided into five sections so that each counted as a separate location (NE, SE, SW, NW, and center), and the other was at a crossroads in Venus, Florida where a BP gas station was on one side of the road and counted as a location and a Circle K gas station was on the other side of the road and counted as a separate location. All BTGR and most GTGR data were collected using a voice recorder (Voice Memos app on an iPhone) and later transcribed into the datasheet in Google Sheets (https://www.google.com/sheets/about/). Some GTGR data were first entered into the program Prim8 Software (https://www.prim8software.com/), and then transcribed into the Google Sheet.
The focal protocol was modified in three ways to accommodate that we were only interested in collecting foraging behavior in BTGR, in comparison to GTGR where we collected data on other behaviors as well. The first way in which the BTGR protocol differed was that we ended each focal when the bird went out of view and started a new focal when they came back in view (rather than allow “out of view” for up to five min per focal). This omitted the problem of trying to account for what might have happened in the time out of view when running the analysis. The second modification was that we did not wait a minimum of three weeks between separate focal follows because we were only looking at food type switch rates and these are, by definition, less autocorrelated with time because a switch could only occur within the same focal follow. This still matches what occurred with the GTGR data because, while the aim was to conduct a focal follow for 10 continuous minutes without the bird going out of view, BTGR were very difficult to follow and they went out of view often and were not findable again for several minutes or even days. In these cases, the observer would continue to seek the bird out for several minutes and then across days to obtain more focal observation time until at least 40 min of focal time per bird across at least four separate days was reached (the equivalent of four 10 min focal follows). Each unique day generally corresponded with the focal number. For example, focal one might have occurred on the first day of observations for that individual, which might have had a few separate bouts of focal observations that added up to 12 min. Focal two on the second day of observations might have only had one bout of focal time that added up to five min. Focal three on the third day might have had nine min of observation time in three bouts. Focal four might have occurred across day four with six minutes of focal time and day five with 12 min of focal time, for an overall total of 44 minutes of observation time across five days. A study on dolphins (Tursiops cf. aduncus) found that focal follow data that was separated by 10.5 min was functionally independent (Karniski et al., 2015). Therefore, changing the three week spacing minimum to one day (until four focal follows – at least 40 min across four separate days – were reached and then spacing them one week apart after) should be enough temporal distance to assume independence of the data points. The third modification was that we collected BTGR data only on the “What” (what did the bird eat, e.g., fries, grains, insect) categories that were found in the GTGR, and we added more food types for the BTGR as needed. We also recorded the latitude and longitude of the location of the grackle at the end of the focal, as in the GTGR protocol.
We set the BTGR minimum sample size to the average number of focal follows obtained from GTGR in Logan et al. (2025): we aimed for a minimum of four focal follows per bird (conducted on separate days between 09:00 and 16:00, attempting to counterbalance mornings and afternoons). The GTGR in the flexibility manipulated condition in Logan et al. (2025) had an average of 3.2 focal follows per individual. Therefore, we used a minimum of four focal follows per BTGR individual (we rounded 3.2 up to the next whole number). When we analyzed food type switching in the two GTGR populations in Logan et al. (2025), we were able to detect differences in food type switching between individuals in the flexibility manipulated (n=6 individuals) and control (n=7 individuals) conditions. Therefore, we set the BTGR minimum sample size at six individuals. This matched with a power analysis in Logan et al. (2025) (in the section: Ability to detect actual effects > Scenario 2) on a very similar model, where we found that we could reliably detect large and medium effects with a sample size of eight in the smallest population. The full experimental protocol can be found in the Supplementary Material (Logan, 2025).
Only those focal follows that contained data on food types, including unknown food types, were included in the analyses. Therefore, focal durations were balanced between the species.
Table 2 - The foraging section of the GTGR ethogram that was used for BTGR data collection. Two new food categories were added that were uniquely seen being eaten by BTGR.
Foraging behavior | Behavior | Description |
WHAT are they eating? | 23 categories | Fry, lizard, unknown, grains (pizza, noodles, bread, rice, chips, rice crispies, crackers, pretzel, muffin, cookie, popcorn), insect, rock, cat food, worm, seed, food crumbs, vegetation (leaf, grass, branch, flower, moss), fruit (flesh, peel), bird poop, candy, vomit, misc. trash (paper, condiment packet), soil (clay, dirt), condiment, carcass, chicken (bone, skin), peanut, mulch. New for BTGR: oil, cheese |
Foraging (state) | Record the start and stop times for each foraging bout | When the bird is searching for food (have to be touching what they are searching through), pecking in the ground, and/or eating food. If the bird pauses foraging behavior for up to 10s, keep this state going. If it pauses for >10s, end the foraging state |
Analyses
A food type switch was counted if a bird ate one food type and then ate another food type. It also counted as a switch if they went back to eating a previous food type. For example, if a bird ate grains, insect, worm, insect, that would count as three switches among three food types. We did not exclude any data (note that there were eight BTGR focal follows that occurred less than a week before the previous focal follow or occurred on the same day as another follow at the same location. However, these focal follows did not contain food type switches and thus were not included in the analyses). When there were missing data (e.g. if a bird did not have any food type or food switching data), then it was not included in the analyses. Following procedures in McElreath (2020), we constructed a hypothesis-appropriate mathematical model for the response variable that examines differences in the response variable between sites. The single population of BTGR in Venus, Florida, was considered a site and we examined differences between BTGR and each of the two GTGR sites. For each focal follow, we calculated the number of switches between food types that occurred and the total amount of time that the bird was observed (using seconds as the unit of time because that was the resolution of data that we collected). We used a rate to be able to directly compare the results among individuals, regardless of any differences in how much time they were followed for. We summed both measures across focal follows to have one data point per bird. This model takes the form of:
𝑠𝑤𝑖𝑡𝑐ℎ𝑒𝑠𝑖 ~ Binomial(𝑡𝑜𝑡𝑎𝑙𝑡𝑖𝑚𝑒𝑖, 𝑝),
logit(𝑝) ~ 𝑎𝑖i[𝑠𝑖𝑡𝑒],
where 𝑠𝑤𝑖𝑡𝑐ℎ𝑒𝑠𝑖 is the sum of the number of times individual, i, changed food types within focal follows that contained food type data, 𝑡𝑜𝑡𝑎𝑙𝑡𝑖𝑚𝑒𝑖 is the number of seconds individual, i, was observed across all of its focal follows that contained food type data, 𝑝 is the probability of switching to a different food type per second, and 𝑎𝑖 is the intercept (average; one per level of 𝑠𝑖𝑡𝑒: GTGR Arizona, GTGR California, and BTGR).
The model was the same for analyzing the variance in switch rates except b[individual] and an extra prior were added as follows:
logit(𝑝) ~ 𝑎𝑖i[𝑠𝑖𝑡𝑒] + b[individual],
which gives the probability of switching food types per bird. We specified the priors for this as:
b[individual] ~ Normal(0, 𝜎[site]),
𝜎[site] ~ Exponential(1),
where 𝜎[site] gives the average variance across individuals per site.
We then performed pairwise contrasts to determine whether there were differences between sites, concluding positively if 89% of the difference between two sites is on the same side of zero (following McElreath, 2020). The Bayesian approach first estimates for each population the most likely distribution of values given the observed sample, and only in a second step do we compare these estimated distributions. The important part here is that the sample size in each population exceeds a given minimum to reliably estimate the distribution for this population. Differences in sample size across populations do not matter with this approach (McElreath, 2020). We ran these analyses in R (current version 4.0.3, R Core Team, 2023) and used the following R packages: rethinking (McElreath, 2020), cmdstanr (Gabry et al., 2025), and dplyr (Wickham et al., 2021). Our code is available at Logan & McCune (2025).
Results
There were no strong differences in the AVERAGE probability of switching among food types per second between BTGR and either population of GTGR, nor between the two GTGR populations (Figure 1; contrasts: GTGR AZ-BTGR: mean difference per second=-0.0004, sd=0.0003, 89%CI=-0.001-0.0001; GTGR CA-BTGR: mean=-0.0002, sd=0.0004, 89%CI=-0.001-0.0004; GTGR AZ-GTGR CA: mean=-0.0002, sd=0.0003, 89%CI=-0.0007-0.0002). This supports Prediction alternative 1.
There were also no strong differences in the VARIANCE of the probability of switching among food types per second between BTGR and either population of GTGR, nor between the two GTGR populations (Figure 1; contrasts: GTGR AZ-BTGR: mean difference per second=-0.16, sd=0.61, 89%CI=-0.99-0.76; GTGR CA-BTGR: mean=0.13, sd=0.73, 89%CI=-0.94-1.28; GTGR AZ-GTGR CA: mean=-0.29, sd=0.45, 89%CI=-0.97-0.37). The variances from the raw data for food switches per minute for each site were 0.01 for BTGR, 0.12 for GTGR Arizona, and 0.14 for GTGR California (note that the Bayesian model accounts for any potential differences in variance because it compares the distributions of data points between all populations). This supports Prediction alternative 1.
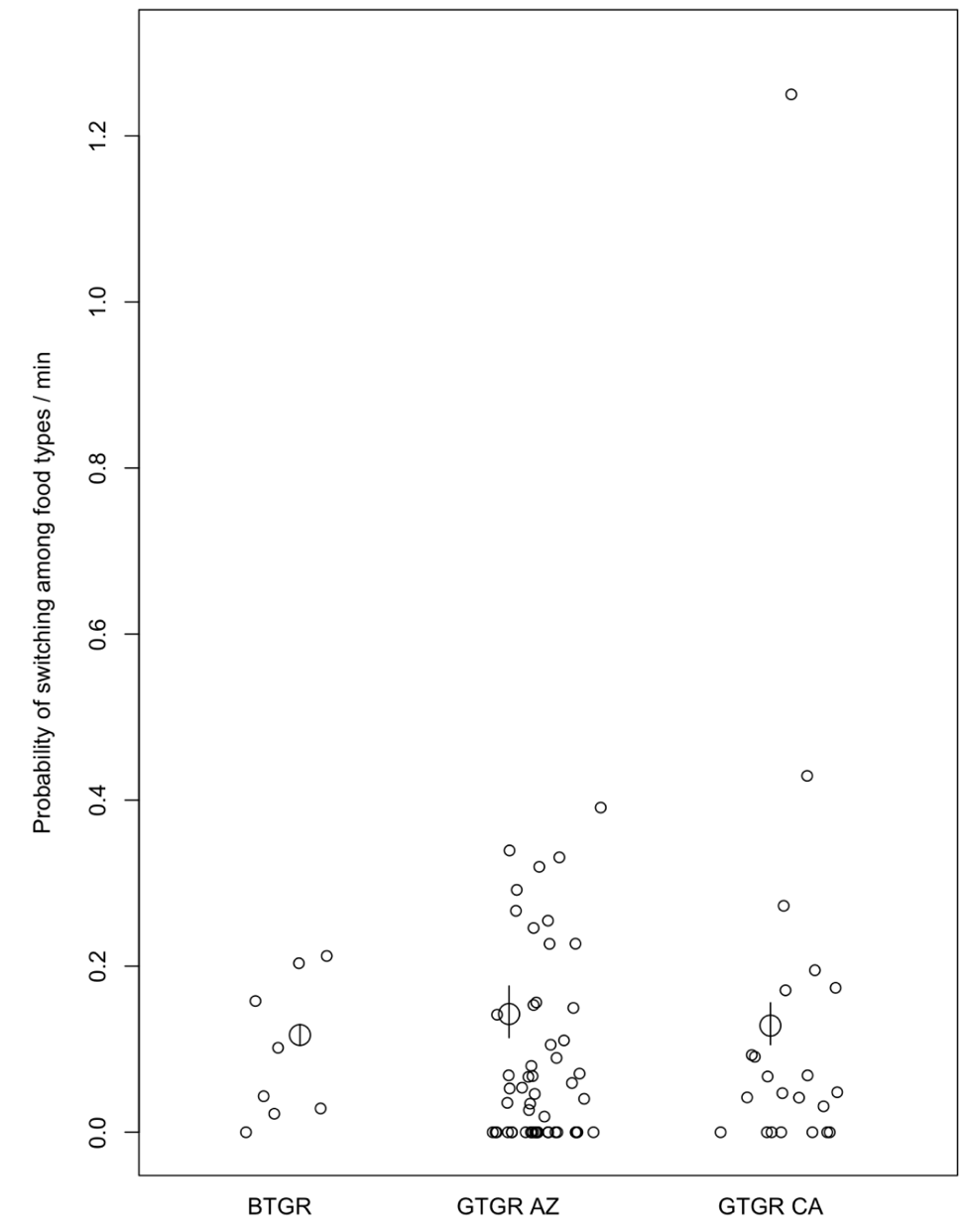
Figure 1 - The probability of switching among food types per minute for the BTGR (n=8), and GTGR in Arizona (n=54) and California (n=22). The small circles are the data points per individual and the large circles are the estimated means with their 89% compatibility intervals represented by the vertical lines.
Discussion
There were no strong differences in flexibility (averages or variances), measured as food type switching during focal follows, between BTGR and GTGR. This converges with the small amount of evidence we were able to obtain from the reversal learning tests in the aviaries from two BTGR individuals: their reversal learning speeds (35 and 79 trials to reverse) were well within the range of the GTGR reversal learning speeds (26-159 trials to reverse; Logan et al., 2023a). We know that GTGR are highly flexible relative to other species (Logan, 2016), therefore the similar levels of flexibility between the two species indicates that BTGR are also highly flexible.
We found no strong differences in flexibility, measured as food type switching, between the two GTGR populations, which supports our finding that there was also no difference in the average flexibility based on reversal learning between these populations (Logan et al., 2025). This provides additional evidence that food type switching is likely measuring the same trait as reversal learning and therefore is a valid measure of flexibility. Logan et al. (2025) did find a difference in the reversal learning flexibility variance between the GTGR populations and, while we found no strong difference in the variance when using food type switching probabilities, the California population, closest to the northern edge of the range, had higher variances, which is in the same direction as what Logan et al. (2023b) found.
Unfortunately, we were not able to obtain comparable exploration, innovativeness, and persistence data on the BTGR to understand whether one or more of these behaviors could relate to species differences in range expansion rates. Before the focal follow data in the current article were collected, we obtained a small sample of data from BTGR in 2022 in aviaries. We reported and analyzed the BTGR 2022 aviary data in a separate article, Logan et al. (2023b), where we analyzed the results from these tests for GTGR, therefore it was a more appropriate place to compare results from similar tests. The analyses suggest that BTGR are less innovative (n=4) and less persistent (n=5) than both GTGR populations, while having similar levels of exploration (n=5) as the California GTGR and being less exploratory than the Arizona GTGR. Indeed, the California GTGR on the northern edge of their range are more persistent than the Arizona GTGR population, though there were no strong differences in innovation or exploration. This lends more support to persistence as the behavior that might play a larger role in the range expansion of the already highly flexible GTGR. However, this is speculative due to the small BTGR aviary sample sizes. More research is needed to make robust comparisons between the two species on these other behaviors.
The lack of a difference in flexibility between the two species suggests that the original behaviors exhibited by these species happen to be suited to the similar challenges of human-modified landscapes where both species are found. This supports the hypothesis that, because these species are closely associated with human-modified environments (Post, 1992; Post et al., 2020; Summers et al., 2023; Wehtje, 2003), which may be similar across the geographic range (what we refer to as “uniform” in the predictions), individuals in new areas may not need to learn much about their new environment. They can eat familiar foods and access these foods in similar ways across their range. For example, food at restaurants and in garbage cans are similar across North America. It is possible that environmental, rather than behavioral, variables play a larger role in restricting the BTGR range expansion. Summers et al. (2023) found that, between 1979 and 2019, BTGR were present in primarily warm, wet, coastal habitats and this did not change over time. The range they expanded into during this time was made suitable to them likely due to climate change. In contrast, GTGR shifted to using more urban, arid habitats over this time, which suggests that behavior could be a key facilitator involved in their range expansion. MacLean & Beissinger (2017) found a similar result using a metaanalysis: range shifts were associated with habitat breadth. Because urban environments represent a rapidly increasing category of environmental change (Goldewijk, 2001; Liu et al., 2020; Wu et al., 2011), our observations highlight the importance of differentiating between whether a behavior might be linked to the ability to live in urban environments versus the ability to expand into new habitats. Flexibility is potentially linked to living in urban environments, but not necessarily to the ability to expand into novel areas.
In conclusion, the evidence that two closely related species have similar levels of flexibility, but different range expansion rates does not support the hypothesis that flexibility is the primary facilitator of rapid geographic range expansions into new areas. This does not rule out that flexibility might be a needed variable to rapidly expand a range, as shown by GTGR who have high average levels of flexibility on and away from the edge, but just because it is present, does not mean that the species will expand their range, as shown by the BTGR. It appears that other behaviors, such as persistence (Logan et al., 2023b), innovation, food neophobia (Liebl & Martin, 2014; Magory Cohen et al., 2020; Martin & Fitzgerald, 2005), exploration (Chuang & Riechert, 2021), and risk aversion (Eccard et al., 2023) may play a more primary role in expanding the edge of the range further. This indicates the importance of investigating multiple behaviors in each species of interest to determine what, if any, role they play and in what direction.
Ethics
This research was carried out in accordance with permits from the:
US Fish and Wildlife Service (scientific collecting permit number MB76700A-0,1,2)
US Geological Survey Bird Banding Laboratory (federal bird banding permit number 23872)
Arizona Game and Fish Department (scientific collecting license number SP594338 [2017], SP606267 [2018], SP639866 [2019], and SP402153 [2020])
Institutional Animal Care and Use Committee at Arizona State University (protocol number 17-1594R)
Institutional Animal Care and Use Committee at the University of California Santa Barbara (protocol number 958)
Institutional Animal Care and Use Committee at Archbold Biological Station (protocol number ABS-AUP-033-R)
California Department of Fish and Wildlife (scientific collecting permit [specific use] number S‐192100001‐19210‐001)
RegionalSan (access permit number AP 2021-01)
Florida Fish and Wildlife Conservation Commission (scientific collecting permit number LSSC-21-00050)
Author Contributions
Logan: Hypothesis development, data collection (BTGR focal follows and field work), data analysis and interpretation, write up, revising/editing, materials/funding.
McCune: Method development, data collection (Arizona and California focal follows and field work), revising/editing.
Rowney: Data collection (Arizona field work), revising/editing.
Lukas: Hypothesis development, data collection (BTGR field work), data analysis and interpretation, write up, revising/editing.
Appendices
Supplementary Material is available at https://doi.org/10.17605/OSF.IO/C9PE8 (Logan, 2025).
Acknowledgements
Preprint version 3 of this article has been peer-reviewed and recommended by Peer Community in Registered Reports (https://doi.org/10.24072/pci.rr.100995; Shafiei Sabet & Mazza, 2025). We thank our Stage 1 manuscript PCI Ecology recommender, Esther Sebastián González, and reviewers Caroline Marie Jeanne Yvonne Nieberding, Pizza Ka Yee Chow, Tim Parker, and 1 anonymous reviewer for their helpful feedback; our Stage 2 reviewers at PCI Ecology, Pizza Ka Yee Chow, Roi Dor, and one anonymous reviewer, and then our PCI Registered Reports recommenders, Valeria Mazza and Saeed Shafiei Sabet, and reviewers, Vedrana Šlipogor and Molly Gilmour for their wonderful support and feedback. Note that ChatGPT was used to figure out how to install two R packages at the Rmarkdown file. We thank Julia Cissewski and Sophie Kaube for tirelessly solving problems involving financial transactions and contracts; and Richard McElreath for generously supporting this project. We are grateful for support for the boat-tailed grackle research: thank you to Sebastian Sosa for advice on how to modify the focal follow protocol for the BTGR; BP gas station and Circle K gas station for allowing research access to their land; Lake Placid Town Hall for allowing research access at Lake June Park; Archbold Biological Station and Buck Island Ranch for allowing access to hundreds of acres for grackle research; Reed Bowman and Sahas Barve for hosting this research at Archbold Biological Station; Betsie Rothermel for consultations during difficult times; Joe Guthrie and Emily Angell for helping protect the aviaries from predators; Brenda Leitner for innovative brainstorming sessions on how to catch grackles; common grackles for enticing the boat-tailed grackles into the trap; and the boat-tailed grackles for participating in this research. We are grateful for support for the GTGR research: thank you to Melissa Wilson for sponsoring our affiliations at Arizona State University and lending lab equipment; Kevin Langergraber for serving as local PI on the ASU IACUC; Kristine Johnson for technical advice on great-tailed grackles; Aaron Blackwell and Ken Kosik for being the UCSB sponsors of the Cooperation Agreement with the Max Planck Institute for Evolutionary Anthropology; Christa LeGrande for conducting focal follows and field work on the California GTGR; Tim Busch at the Woodland-Davis Clean Water Agency, Bryan Young and Chris Conard at RegionalSan, and Conaway Ranch for hosting the California field research on their land; Luisa Bergeron, Zoe Johnson-Ulrich, and Melissa Folsom for conducting focal follows and field research, August Sevchik for helping in the field and in the aviaries, and Sawyer Lung for field support in Arizona; our Arizona research assistants: Aelin Mayer, Nancy Rodriguez, Brianna Thomas, Aldora Messinger, Elysia Mamola, Michael Guillen, Rita Barakat, Adriana Boderash, Olateju Ojekunle, August Sevchik, Justin Huynh, Jennifer Berens, Amanda Overholt, Michael Pickett, Sam Munoz, Sam Bowser, Emily Blackwell, Kaylee Delcid, Sofija Savic, Brynna Hood, Sierra Planck, and Elise Lange; and the great-tailed grackles for participating in this research.
Funding
This research was funded by the Department of Human Behavior, Ecology and Culture at the Max Planck Institute for Evolutionary Anthropology.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article. The authors declare the following non-financial conflict of interest: Logan and Lukas are Recommenders at PCI Ecology and PCI Registered Reports, and Logan was on the Managing Board at PCI Ecology (2018-2022) and is on the Managing Board at PCI Registered Reports (2021-current).
Data, scripts, code, and supplementary information availability
Data and code are available online: https://doi.org/10.5063/F18P5Z09 (Logan & McCune, 2025). Supplementary information is available online: https://doi.org/10.17605/OSF.IO/C9PE8 (Logan, 2025).