
 CC-BY 4.0
CC-BY 4.0
Introduction
The family Scombridae includes mostly epipelagic marine fishes, such as tunas, a large, epipelagic predator (Collette & Nauen, 1983). The four genera of tuna, Auxis, Euthynnus, Katsuwonus, and Thunnus, form the tribe Thunnini. Among the Thunnini, the genus Auxis is an epipelagic, neritic, and oceanic genus found worldwide in tropical and subtropical oceans (Collette & Nauen, 1983). Auxis consumes various fishes, crustaceans, cephalopods, and other prey and is preyed upon by large tunas, billfishes, barracudas, sharks, and more (Collette & Nauen, 1983). Auxis comprises two extant species: the frigate and bullet tunas (Auxis thazard and Auxis rochei). They exhibit significant morphological similarities (Vieira et al., 2022) and little osteological differences. The fossil history of Auxis is incomplete due to a lack of fossil records and many invalidations of fossil specimens. The fossilization potential is extremely low for taxa in deep-sea and pelagic environments with a 15% and 3% possibility of fossilization respectively due to factors like the fragile skeleton and high environmental stress (Shaw et al., 2020). As a result, Auxis fossils are extremely rare, with very few reported specimens. Meanwhile, several fossil specimens previously identified as Auxis have undergone multiple taxonomic changes within the Scombridae (Nam et al., 2021). Several cases of Auxis misidentification have been noted in the literature. Five scombrid species from the Miocene in Europe were initially identified as Auxis by Kramberger-Gorjanović (1882) and Gorjanović-Kramberger (1895); however, Nam et al. (2021) later disputed these identifications based on differences in traits such as vertebral count, lack of the haemal arch, and gill cover and dentition size. Additionally, Woodward (1901) reclassified a scombrid fossil originally described by Agassiz (1833–1844) as Auxis, representing what was considered the earliest record of Auxis from the Eocene. Bannikov and Sorbini (1984) later corrected this identification, noting discrepancies in vertebral number and shape of haemal arches. Another specimen, a Middle to Upper Miocene scombrid from Russia identified as Auxis by Bogatshov (1933), was reclassified by Bannikov (1985) due to an inconsistent vertebral count. With these invalidations, the only currently accepted fossil record of Auxis dates back to the Miocene and is reported from the same formation as the specimen described in this paper (†Auxis koreanus, Nam et al., 2021). Moreover, the detailed study of the vertebral anatomy of Auxis has been hindered by the paucity of recovered specimens including both skulls and vertebrae.
An imprint of tuna vertebrae was collected from the Duho Formation, Pohang City, South Korea, in 2020 (Figure 1). The new specimen (GNUE322001, Gongju National University of Education) represents the second discovery of Auxis from the Duho Formation of the Korean Peninsula and the second valid Auxis specimen in the world. Although the specimen is preserved poorly and lacks cranial elements, it possesses diagnostic characters of the vertebrae of the genus Auxis: the bifurcated inferior antero-zygapophysis with a long pedicle and no trellis. This paper describes this new tuna specimen and discusses the palaeoecological implications of the presence of tunas in the Miocene of South Korea.
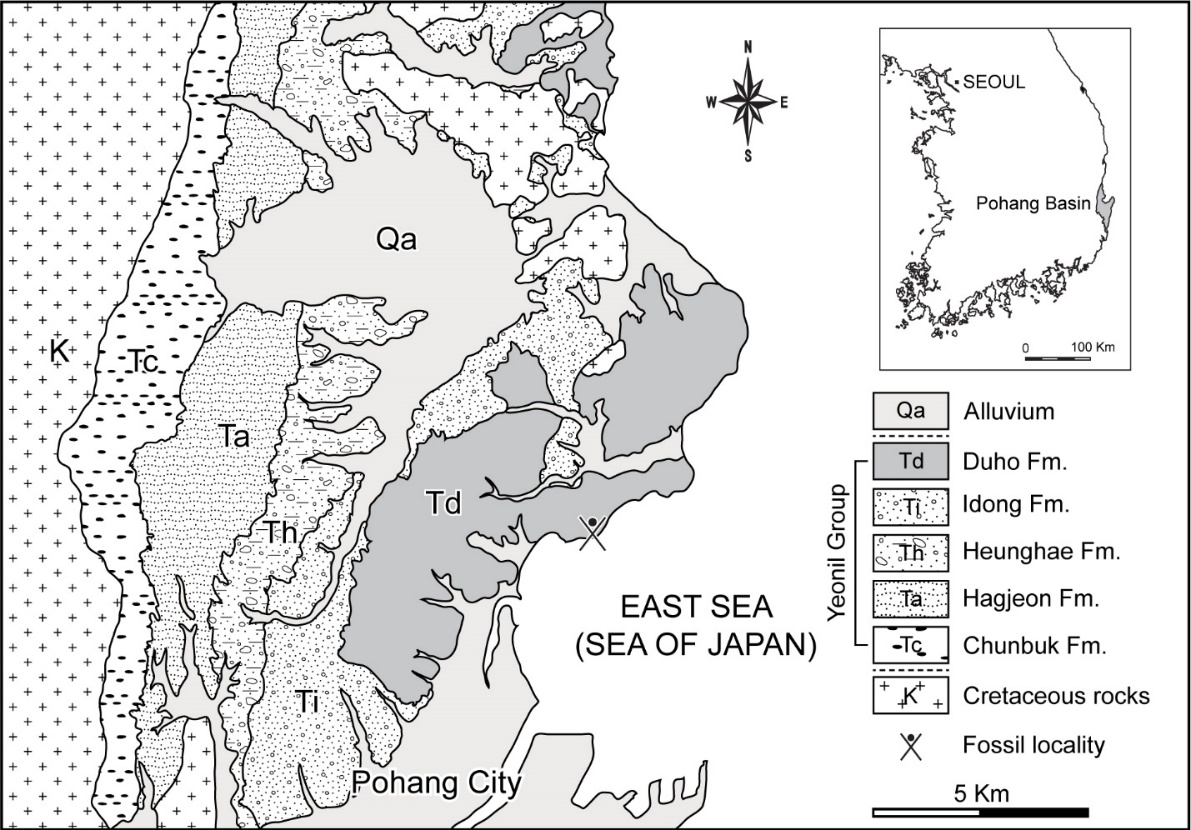
Figure 1 - Geologic map of the northern part of the Pohang area with Cenozoic basins in South Korea (inset), depicting the fossil locality where GNUE322001 was collected.
Geological setting
The Pohang Basin is the largest Cenozoic basin in South Korea (Yoon, 1975; Figure 1) and is a pull-apart basin that started to form by post-volcanism subsidence at ~17 Ma (Sohn et al., 2001). The Yeonil Group, in the Pohang Basin, includes more than 1 km thick non-marine to deep-marine strata that are characterized predominantly of clastic sediments of marine origin (Sohn et al., 2001; Kim, 2008). This group comprises conglomerates and sandstones along the basin margin and hemipelagic mudstones and sandstones towards the basin center (Sohn et al., 2001; Woo & Khim, 2006). The Yeonil Group consists of the Duho, Idong, Heunghae, Hagjeon, and Chunbuk Formations (Yoon, 1975; Yun, 1986; Figure 1). The Duho Formation, where the studied specimen was collected, occurs in the uppermost part of the Yeonil Group and is about 250 m thick (Yun, 1986). A pale grey to light brown homogeneous mudstone with intercalated sandstone is the main deposit of the Duho Formation (Hwang et al., 1995; Kim & Paik, 2013). The Duho Formation produces a variety of marine invertebrate and vertebrate fossils, including mollusks (Kim & Lee, 2011; Kong & Lee, 2012), fishes (Ko, 2016; Ko & Nam, 2016; Kim et al., 2018; Nam et al., 2019; Nam et al., 2021; Nam & Nazarkin, 2022; Nazarkin & Nam, 2022; Malyshkina et al., 2023; Table 1), and whales (Lim, 2005; Lee et al., 2012). Such a diverse fossil record has produced equally diverse paleoenvironmental interpretations of the depositional environment of the Duho Formation. The paleoenvironmental interpretation of the Duho Formation ranges between shallow marine (Kim, 1965; Yun, 1985), offshore (Yoon, 1975, 1976; Lee, 1992), low energy (Seong et al., 2009; Kim & Lee, 2011), hemipelagic (Chough et al., 1990; Kim & Paik, 2013), and deep-sea environments (Chough et al., 1990; Kim & Paik, 2013). Various studies on the age of the Duho Formation additionally resulted in diverse interpretations (Kim et al., 2018), ranging from the Early Miocene based on Zircon dating (Lee et al., 2014), Middle Miocene based on paleomagnetic dating and volcanic rocks (Kim et al., 1993; Chung & Koh, 2005), and Late Miocene based on dinoflagellate and radiolarian fossils (Byun & Yun, 1992; Bak et al., 1996).
Table 1 - List of fish taxa from the Duho Formation.
Taxa | References | |
|---|---|---|
Actinopterygii | Pleuronectiformes | |
Pleuronichthys sp. | ||
†Vinciguerria orientalis | ||
†Stenobrachius sangsunii | ||
Vinciguerria sp. | ||
†Auxis koreanus | ||
†Carcharodon hastalis | ||
Elasmobranchii | Hexanchus griseus | |
†Dalatias orientalis | ||
Mitsukurina owstoni | ||
†Otodus megalodon | ||
†Parotodus benedenii | ||
†‘Isurus’ planus | ||
Isurus sp. | ||
†Cetorhinus huddlestoni | ||
Carcharhinus aff. C. plumbeus | ||
Carcharhinus aff. C. amblyrhynchos | ||
Carcharhinus aff. C. altimus | ||
†Galeocerdo aduncus | ||
Materials and methods
The specimen GNUE322001, a partially preserved caudal tuna vertebrae imprint, is housed in the Gongju National University of Education (GNUE), Gongju City, South Korea. The specimen was photographed using a digital camera (Sony A7R4A). Image processing and line drawings of the specimen were done using Adobe Photoshop v 23.4.2. and Adobe Illustrator v 26.4.1. All measurements were taken using a digital caliper.
Anatomical nomenclature
We follow the terminology of Starks (1910), which was applied to Auxis, to describe peculiar vertebral structures of the studied specimen and occasionally refer to the terminology of Romeo and Mansueti (1962) for efficient comparison between Auxis, Euthynnus, and Katsuwonus of the tribe Thunnini.
Repositories and institutional abbreviation
The specimen is deposited in the Gongju National University of Education (GNUE), Gongju City, South Korea.
Results
Systematic Paleontology
Order Perciformes Nelson, 2006
Suborder Scombroidei Nelson, 2006
Family Scombridae Rafinesque, 1815
Tribe Thunnini Starks, 1910
Genus Auxis Cuvier, 1829
Auxis sp.
Type species
Scomber rochei Risso, 1810
Occurrence
Duho Formation, Hwanho-dong, Buk-gu, Pohang City, North Gyeongsang Province, South Korea (N36°3’49.10”, E129°23’47.07”) (Figure 1), preserved in a massive grey mudstone in the Duho Formation (Figure 2).
Description
Due to the dissolution of the original bones, only the molds of the eight articulated caudal vertebrae are partially preserved (Figure 2). In particular, due to the breakage of the matrix, only small fragments of the first and last vertebrae are preserved. The centra have an amphicoelous shape, consisting of two robust cones, and both cones are connected by what appears as a wide foramen. In all Thunnini and certain fish vertebrae, the centrum is not pierced thus lacks a notochordal foramen (Starks, 1910; Graham & Dickson, 2000). Therefore, the apparent wide foramen in GNUE322001 is interpreted as the result of the split of the specimen along a parasagittal plane, and does not represent a real anatomical trait. The anteroposterior length and dorsoventral height of the centrum are subequal, and the dorsal and ventral margins of the centrum are slightly concave in lateral view.
The superior antero-zygapophysis is quite large and dorsoventrally deep, covering most of the posterodorsal margin of the preceding centrum from the posterior margin of the centrum to the posterior edge of the base of its dorsal spine (Figure 2). In contrast, the superior postero-zygapophysis is weakly developed and is barely discernable in lateral view due to the overlapping superior antero-zygapophysis of the following vertebra.
The dorsal spine originates from the centrum at mid-length, and is slightly angled posteriorly, forming an angle of ~80-85° with the posterodorsal margin of the centrum (Figure 2). It slightly curves posteriorly at a third of the total length of the preserved spine from its base.
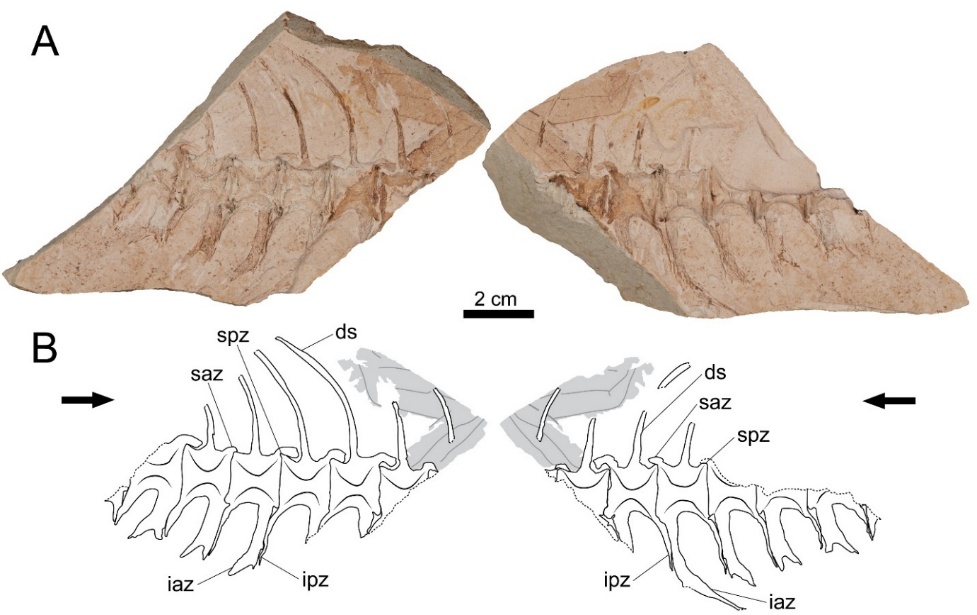
Figure 2 - Photographs and drawings of GNUE322001. (A) Photographs of GNUE322001. Each counterpart mold shows a lateral side of the vertebrae without the original bones. (B) Drawings of GNUE322001. Black arrows point towards the anterior direction of the vertebrae. Dashed lines indicate a broken edge. Grey areas indicate an unidentified leaf imprint. Vertebral terminology follows Starks (1910). Abbreviations: ds, dorsal spine; iaz, inferior antero-zygapophysis; ipz, inferior postero-zygapophysis; saz, superior antero-zygapophysis; spz, superior postero-zygapophysis.
On the fourth to seventh vertebrae, the preserved inferior antero- and postero-zygapophyses project from the centrum ventroposteriorly at an angle of ~70-80° (Figure 2). The length of these ventral processes of the vertebrae progressively decreases in more posterior vertebral positions. The length of these processes in the first to third vertebrae cannot be assessed due to incomplete preservation.
All preserved inferior antero-zygapophyses are bifurcated into anterior and posterior branches, and the latter tends to be longer (Figure 2). The inferior antero-zygapophysis of the fourth vertebra is much longer than that of the other vertebrae. It extends nearly to the level of the posterior tip of that of the following vertebra. The inferior postero-zygapophysis extends almost to the ventral tip of the anterior branch of the inferior antero-zygapophysis of the following vertebra. They firmly attach to each other along the entire posterior margin of the inferior postero-zygapophysis.
Remarks
We classify GNUE322001 as anterior to the middle caudal vertebrae based on the progressively decreasing lengths of the ventral processes, a pattern also observed in the caudal vertebrae of extant Auxis (see Uchida, 1981: figure 24; Jawad et al., 2013: figure 1; Figure 4).
The classification of extant Auxis is based primarily on the relative body depth, corset width, the number of gill rankers and color pattern (Collette & Aadland, 1996). The extinct Auxis, †A. koreanus, is distinguished from extant Auxis by the osteological differences in skull elements (Nam et al., 2021). Because only the caudal vertebrae are preserved in GNUE322001, the skull is not available for comparison with other species of Auxis. However, GNUE322001 exhibits several distinct morphological features in the caudal vertebrae, which we compare with that of other extant Auxis species in the following discussion.
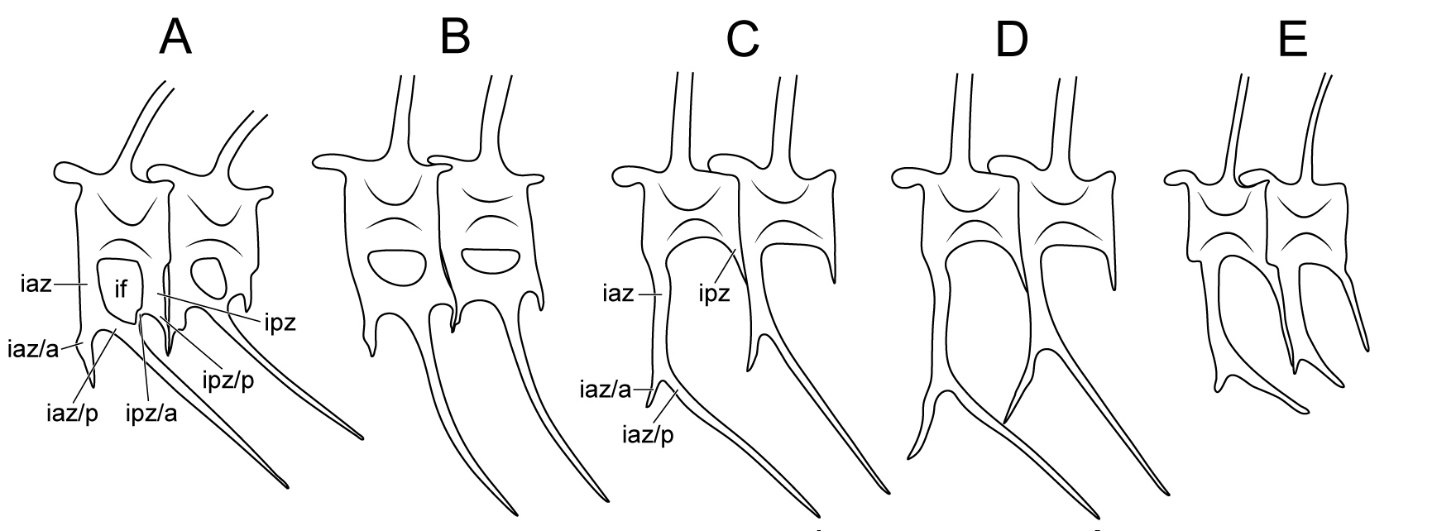
Figure 3 - Comparative diagram of the anterior caudal vertebrae of Auxis, Euthynnus, Katsuwonus, and GNUE322001. (A) Euthynnus. (B) Katsuwonus. (C) A. rochei. (D) A. thazard. (E) GNUE322001 (Godsil & Byers, 1944; Yoshida, 1979; Uchida, 1981). Vertebral terminology follows Starks (1910). Abbreviations: iaz/a, anterior branch of inferior antero-zygapophysis; iaz/p, posterior branch of inferior antero-zygapophysis; ipz/a, anterior branch of inferior postero-zygapophysis; ipz/p, posterior branch of inferior postero-zygapophysis; if, inferior foramen. Note that in Euthynnus (A) and Katsuwonus (B), the posterior branch of inferior antero-zygapophysis and the anterior branch of inferior postero-zygapophysis are fused, forming the inferior foramen and trellis. Size not to scale.
Discussion
Anatomical comparisons
Among the Thunnini, the genera Auxis, Euthynnus, and Katsuwonus share a morphological similarity in the inferior antero-zygapophysis in that it is bifurcated into anterior and posterior branches, a unique characteristic only observed in these three genera. However, Auxis exhibits ventral bifurcation only in the caudal vertebrae, whereas this trait is also found in the posterior abdominal vertebrae in Euthynnus and Katsuwonus (see Godsil & Byers, 1944: figure 19; Godsil, 1954: figure 83; Yoshida & Nakamura, 1965: figure 3). Furthermore, the pedicle of Auxis, a median rod formed by the fusion of both sides of the inferior antero-zygapophyses below the centrum and above the haemal canal (Kishinouye, 1923), is far longer than that of Euthynnus and Katsuwonus (Godsil, 1954; Figure 3).
Most significantly, the anterior caudal vertebrae in Euthynnus and Katsuwonus are characterized by the presence of an inferior foramen and trellis pattern. The inferior foramen is a hole-like structure fenestra created by the complete fusion of the posterior branch of the inferior antero-zygapophysis (prehaemapophysis of Romeo & Mansueti, 1962) and the anterior branch of the inferior postero-zygapophysis (posthaemapophysis of Romeo & Mansueti, 1962) under the centrum (see Romeo & Mansueti, 1962: figure 2D; Figure 3A, B). The trellis pattern is formed by the continuous repetition of the inferior foramen across the vertebrae (Figure 3A, B). In Auxis, the inferior foramen and trellis pattern are scarcely developed, and even when present, only occur in the posterior caudal vertebrae, unlike in Euthynnus and Katsuwonus (Kishinouye, 1923; Godsil, 1954; Figure 3).
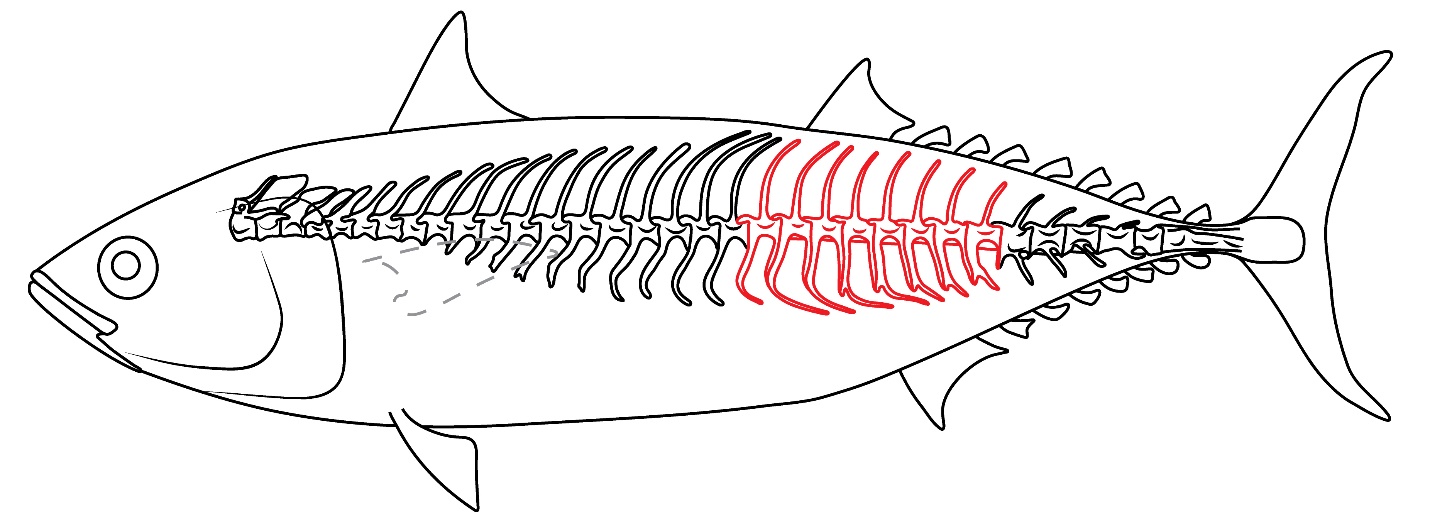
Figure 4 - Reconstruction of GNUE322001. The red-lined vertebrae indicate the possible position of the vertebrae of GNUE322001 in the vertebral column.
Although the cranial elements are not preserved in GNUE322001, this specimen was identified as Auxis primarily based on having a bifurcated inferior antero-zygapophysis with a long pedicle and no trellis. Based on the vertebral column of extant Auxis (see Uchida, 1981: figure 24; Jawad et al., 2013: figure 1), it is suggested that GNUE322001 represents the anterior to the middle caudal vertebral series (Figure 4) as indicated by the lengths of ventral processes that progressively decrease throughout the vertebral series in this taxon.
There are three valid taxa within Auxis, including an extinct species (A. thazard, A. rochei, and †A. koreanus) (Collette & Aadland, 1996; Nam et al., 2021). GNUE322001 is morphologically similar to the vertebrae of A. rochei in that the anterior branch of the inferior antero-zygapophysis is short and does not reach the preceding inferior antero-zygapophysis (Yoshida & Nakamura, 1965; Uchida, 1981; Figure 3C, E). In A. thazard, the anterior branches of the inferior antero-zygapophyses are long enough to contact the preceding inferior antero-zygapophyses (Figure 3D). Comparisons with the extinct taxon †A. koreanus are not possible because the caudal vertebrae are unknown in this species, as they are not preserved in its only known specimens (Nam et al., 2021). Although †A. koreanus was also discovered in the Duho Formation, as was GNUE322001, it is challenging to assign GNUE322001 to †A. koreanus based solely on their shared occurrence within the same formation. Furthermore, the centra of †A. koreanus and GNUE322001 differ significantly in size, with lengths of approximately 0.5 and 1.5 cm respectively (Nam et al., 2021; Figure 2). Therefore, additional study and discovery of Auxis specimens from the Duho Formation are necessary to determine the relationship between GNUE322001 and †A. koreanus, as well as other extant species of Auxis.
Paleoenvironmental perspectives
The major opening of the East Sea that occurred between 23 and 18 Ma widened the gap between the Japanese Arc and the Korean Peninsula by 200-250 km (Sohn et al., 2001). The opening of the East Sea would have facilitated the creation of a variety of marine environments. This increased environmental complexity likely provided habitats that were able to support a wider range of marine species. A diverse fossil record of large oceanic animals in the East Sea during this period such as the tunas (Nam et al., 2021; GNUE322001 in this paper), sharks (Kim et al., 2018), and whales (Lim, 2005; Lee et al., 2012) aligns with this theory.
Upwelling regions, although only constituting 0.1% of the total ocean areas (Wang & Lee, 2019), are where fishes are most abundant due to high production rates (Lalli & Parsons, 1997). One of such fishes is the tuna, which are attracted by the zones of foraging availability created by upwelling zones (Grandperrin, 1978; Nicol et al., 2014). Additionally, based on the record of the fossilized diatom resting spores, which indicate an upwelling activity in the Duho Formation (Hargraves, 1979; Lee, 1993), Kim et al. (2018) hypothesized that the biodiversity of the East Sea increased due to the influence of upwelling during the deposition of the Duho Formation. Thus, it can be concluded that upwelling activity during the Miocene increased pelagic fishes’ and their preys’ biodiversity in the East Sea.
Today, tunas inhabit tropical and subtropical epipelagic ocean ecosystems (Collette & Nauen, 1983). During warm seasons, tunas move into coastal areas, whereas in colder seasons, they exclusively occupy deeper offshore waters (Kishinouye, 1923). In deep epipelagic environments, tunas specifically inhabit deep rocky banks and forage in the deep scattering layer (Kishinouye, 1923; Graham & Dickson, 2000; Graham & Dickson, 2004). Graham and Dickson (2004) argue that most oceanic physical and biological features observed in ocean ecosystems today have been in place since the Miocene. This suggests that the East Sea during the Miocene likely supported a pelagic and subtropical environment, as tuna, which inhabit such ecosystems, were present. This interpretation is supported by researchers who consider the Duho Formation to represent a deep-sea and subtropical environment, among the various interpretations of its environmental context (see the Geological Setting section). Recent discoveries of pelagic sharks (Malyshkina et al., 2023), lightfishes (Nam et al., 2019; Nazarkin & Nam, 2022), and lanternfish (Nam & Nazarkin, 2022) provide evidence that the Duho Formation represents a deep-sea environment. Moreover, based on discoveries of certain plant fossils, some researchers interpret the Duho Formation to have been a subtropical environment (Kim, 2008; Jung & Lee, 2009; Kim et al., 2009; Kim et al., 2017). The discovery of GNUE322001 supports both of these interpretations.
Taphonomic interpretations
The absence of anal pterygiophores in GNUE322001, which in tunas are located directly under the prehaemapophyses (Figure 2), suggests that the specimen underwent significant decomposition underwater. The first steps of decomposition of a fish involve the disarticulation of the jaw and external scales as soft tissues (muscles, skins) decompose (Burrow & Turner, 2012). However, body parts are often disarticulated but still loosely connected (Burrow & Turner, 2012). At this stage, invertebrate and vertebrate scavengers completely disconnect the bones by feeding on the soft tissue or the bones themselves (Burrow & Turner, 2012). In GNUE322001, the absent anal pterygiophores would have been disconnected and/or consumed by marine scavengers, indicating that the vertebrae would have been underwater for a long time. However, the exact taphonomic time frame cannot be determined with the partially preserved vertebrae.
An unidentified leaf imprint is preserved on the anterior portion of the vertebrae of GNUE322001 (Figure 2). Since the fine-grained matrix indicates that the specimen was buried in a low-energy sedimentary environment at the deep-sea bottom, the leaf associated with GNUE322001 would have traveled from shore to the depths of the sea. The leaf exhibits tears on its edges, a characteristic of the fragmentation stage of decomposition where marine detritivorous invertebrates feed on deposited leaves (Bridgham & Lamberti, 2009). The decomposition rate during fragmentation varies depending on salinity; aquatic ecosystems with lower salinity are correlated with faster decomposition (Quintino et al., 2009). Thus, decay rates are greatest in freshwater ecosystems, followed by transitional communities, and slowest in marine ecosystems (Quintino et al., 2009). While the torn edges of the leaf imprint associated with GNUE322001 resemble those of leaves that underwent a two-week decomposition in transitional communities (Bridgham & Lamberti, 2009: figure 15.2), leaves deposited in marine ecosystems take more than twice the time to exhibit a similar amount of biomass remain (Quintino et al., 2009: figure 4). Thus the leaf associated with GNUE322001 would have decomposed after a month of being exposed to water. Although the vertebrae and leaf have experienced different decompositions in isolated conditions, based on the taphonomic time frame inferred from the preservation of the leaf imprint, it can be estimated that the decomposition of GNUE322001 took at least a month. However, perfectly preserved leaves were also reported from the Duho Formation (Jung & Lee, 2009); therefore, the taphonomic scenario inferred from GNUE322001 does not represent a general depositional condition of the Duho Formation.
Acknowledgments
We thank Professor Yuong-Nam Lee for improving the quality of this manuscript. We also thank two anonymous reviewers for their valuable feedback and Dr. Adriana López-Arbarello for overseeing the review process and providing valuable suggestions. Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Paleontology (https://doi.org/10.24072/pci.paleo.100361; López-Arbarello, 2025).
Funding
This work was not supported by any funding source.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Author contributions
Dayun Suh contributed to conceptualization, formal analysis, investigation, visualization, writing of the original draft, and writing of review and editing. Su-Hwan Kim contributed to conceptualization, formal analysis, investigation, methodology, supervision, validation, visualization, and writing of review and editing. Gi-Soo Nam contributed to resources, and validation, and writing of review and editing.