
 CC-BY 4.0
CC-BY 4.0
Introduction
Bacteria of the genus Rickettsiella (order: Legionellales, family: Coxiellaceae) are emerging as both widespread and genetically diverse in arthropods (Leclerque et al. 2011, Mathew et al. 2012, Leclerque 2008, Duron et al. 2017, Duron et al. 2015a, Yaripour et al. 2024, Akter et al. 2024). Historically, Rickettsiella have been identified as contagious entomopathogens infecting a broad range of insects, but various pathotypes have also been reported in other arthropods, including arachnids and crustaceans (Leclerque et al. 2011, Mathew et al. 2012, Leclerque 2008, Cordaux et al. 2007, Zchori-Fein & Bourtzis 2011, Crotti et al. 2010, Bouchon et al. 2016). In infected arthropods, Rickettsiella-induced diseases often replace body cavity tissue with a white iridescent liquid, leading to the death of hosts (Zchori-Fein & Bourtzis 2011, Crotti et al. 2010, Bouchon et al. 2016), which has spurred research into using pathogenic Rickettsiella as a biocidal agent (Zchori-Fein & Bourtzis 2011, Ruiu et al. 2013). However, further surveys have revealed that non-pathogenic Rickettsiella are common in field populations of arthropods (Tsuchida et al. 2014, Nikoh et al. 2018, Tsuchida et al. 2010, Iasur-Kruh et al. 2013, Gauthier et al. 2015). Most of these non-pathogenic Rickettsiella are predominantly maternally transmitted, and have a variety of repercussions on the arthropod’s phenotypes, ranging from providing fitness advantages to manipulating reproduction (Tsuchida et al. 2010, Łukasik et al. 2013, Rosenwald et al. 2020, Polin et al. 2015). In aphids, Rickettsiella viridis can alter body colour, reducing attractiveness to predators and parasitoids (Tsuchida et al. 2010, Polin et al. 2015), or directly protect from entomopathogenic fungi, significantly lowering aphid mortality (Tsuchida et al. 2010, Łukasik et al. 2013). In the poultry red mite Dermanyssus gallinae, Rickettsiella may be an essential nutritional endosymbiont providing B vitamins that are lacking in the blood diet of its host (Price et al. 2021). In the agricultural spider Mermessus fradeorum, a Rickettsiella endosymbiont induces cytoplasmic incompatibility (CI), that results in embryonic mortality in the offspring of matings between infected males and uninfected females (Rosenwald et al. 2020). This reproductive manipulation reduces the production of uninfected progeny and increases the proportion of infected females (i.e., the transmitting sex), thereby promoting the spread of CI-inducing bacteria within the host population (Engelstädter & Hurst 2009). Overall, members of the genus Rickettsiella have evolved complex lifestyles that can significantly impact the ecology and evolution of their arthropod hosts (Duron et al. 2018).
Rickettsiella is remarkably common in the two main tick families, Ixodidae (hard ticks) and Argasidae (soft ticks), with at least 15% of species infected and prevalence ranging from 5% to 100% at the population level (Duron et al. 2017, Duron et al. 2015a, Duron et al. 2016, Garcia-Vozmediano et al. 2022, Vilcins et al. 2009, Anstead & Chilton 2014, Carpi et al. 2011, Kurtti et al. 2002, Leclerque & Kleespies 2012). Rickettsiella also exhibit significant genetic diversity both within and between tick species (Duron et al. 2017, Duron et al. 2015a, Duron et al. 2016, Garcia-Vozmediano et al. 2022, Vilcins et al. 2009, Anstead & Chilton 2014, Carpi et al. 2011, Kurtti et al. 2002, Leclerque & Kleespies 2012). Indeed, genetically distinct strains of Rickettsiella were detected in populations of the castor bean tick Ixodes ricinus and the polar seabird tick Ixodes uriae (Duron et al. 2016, Garcia-Vozmediano et al. 2022). The Rickettsiella-induced phenotypes in ticks have not been clearly identified yet, but a Rickettsiella endosymbiont was observed in a parthenogenetic laboratory colony of the wood tick Ixodes woodi (Kurtti et al. 2002, Leclerque & Kleespies 2012). Both males and females are normally present in this tick species, hence Rickettsiella might manipulate the tick reproduction to induce asexuality (Kurtti et al. 2002). A bacterium was identified in I. ricinus, and further in the blacklegged tick Ixodes scapularis (Merenstein et al. 2020) as a new genus and species, Diplorickettsia massiliensis (Mediannikov et al. 2010), but it was further synonymised as Rickettsiella massiliensis (Saini & Gupta 2021) since the bacterium is nested within the current Rickettsiella genus (Leclerque & Kleespies 2012). Rickettsiella massiliensis has been proposed as a potential agent of opportunistic infections in humans (Subramanian et al. 2011), but no clear clinical cases have been reported to date.
Rickettsiella is currently among the lesser-known tick-borne bacteria (Duron et al. 2018), whereas other Legionellales, particularly those that infect humans, are better understood. The order Legionellales includes two families, Legionellaceae and Coxiellaceae, with several clinically significant representatives, such as Legionella pneumophila, the causative agent of Legionnaires’ disease, and Coxiella burnetii, responsible for Q fever, both of which have been the focus of extensive genomic studies (Duron et al. 2018, Duron et al. 2015b). These pathogenic Legionellales have evolved specific mechanisms to infect mammalian alveolar macrophages, such as a Dot/Icm type IV secretion system to translocate effectors and inhibit host cell apoptosis (van Schaik et al. 2013, Isberg et al. 2008). However, not all Legionellales are pathogens: Some are B vitamin provisioning endosymbionts essential for the nutrition of arthropods with an obligate hematophagous lifestyle (Price et al. 2021, Duron et al. 2018, Duron et al. 2015a, Duron & Gottlieb 2020, Tamarit 2023), including Coxiella-like endosymbionts in ticks (Gottlieb et al. 2015, Santos-Garcia et al. 2023, Guizzo et al. 2017) and Legionella polyplacis in rat lice (Ríhová et al. 2017). Blood is nutritionally unbalanced, and these obligate hematophagous parasites have evolved narrow associations with B vitamin provisioning symbionts, as they themselves cannot synthesise the essential cofactors and vitamins that are lacking in their diet (Duron & Gottlieb 2020, Tamarit 2023). Additionally, Coxiella-like endosymbionts produce chorismate, a tryptophan precursor that regulates serotonin biosynthesis, actively promoting tick feeding activity and blood intake, and modifying behaviour (Zhong et al. 2021).
Compared to other Legionellales, few Rickettsiella genomes have been sequenced. They have circular 1.3-1.9 Mb genomes, smaller than the genomes of other Legionellales such as the pathogens L. pneumophila (3.4 Mb) and C. burnetii (2.0 Mb), and the members of the genera Aquicella (2.5 Mb) and Berkiella (3 - 3.6 Mb), associated with amoebae living in aquatic environments (Duron et al. 2018, Saini & Gupta 2021, Graells et al. 2018). While Rickettsiella genomes are known to encode 1,400-2,200 protein-coding genes (Mathew et al. 2012, Price et al. 2021, Nikoh et al. 2018, Wang & Chandler 2016), no comparative studies have been conducted. So far, the genomes have been used in phylogenomic studies (Saini & Gupta 2021, Santos-Garcia et al. 2023), and a few investigations have been conducted focusing on single strains. Indeed, the Rickettsiella genome of the poultry red mite D. gallinae contains more than 300 pseudogenized protein-coding sequences, but has retained several B vitamin biosynthesis pathways, suggesting the importance of these pathways in the evolution of a nutritional endosymbiosis (Price et al. 2021). Other examples include the genomes of R. grylli, infecting isopods, and R. viridis, infecting aphids, which both contain a complete Dot/Icm type IV secretion system (Nikoh et al. 2018, Wang & Chandler 2016). Previous studies have pointed at R. viridis as being a mutualist (Tsuchida et al. 2014) and at the Rickettsiella isolated from pillbugs as being present in symptomatic (R. grylli, Dittmer et al. 2016), but also in asymptomatic individuals (potentially a different strain, and in coinfection with R. grylli, Bouchon et al. 2016). However, many phenotypically characterised Rickettsiella strains, as the one inducing CI in spiders (Rosenwald et al. 2020), have never been sequenced. Among tick-borne Rickettsiella, only R. massiliensis has had its genome sequenced (Mathew et al. 2012).
In this study, we present five new genomes of Rickettsiella naturally infecting ticks, retrace their evolutionary history, characterise their genomic content, and infer their metabolic capacities. To this aim, we sequenced five microbial metagenomes, including three from I. ricinus, one from Ornithodoros erraticus and one from Ornithodoros phacochoerus. We then compared these genomes with published genomes of Rickettsiella and representative Legionellales, either pathogenic (L. pneumophila, C. burnetii) or symbiotic (Coxiella-like endosymbionts of ticks), to assess gene content and metabolic similarities. We further used these genomic analyses to infer how Rickettsiella can affect the phenotype of their arthropod hosts.
Methods
Sample collection, DNA extraction and sequencing
Specimens of three tick species, I. ricinus, O. erraticus and O. phacochoerus, were collected in this study (see Table 1 for details). No data on sex ratio of these tick species were collected in this article nor in previous studies. Different methods for DNA extraction and sequencing were applied depending on tick species. In all cases, total DNA (tDNA) was extracted from the obtained pellet using the DNeasy Blood and Tissue Kit (Qiagen). The obtained tDNA was quantified on Qubit using the dsDNA high-sensitivity kit (Invitrogen). Finally, a shotgun metagenomics approach was applied, obtaining reads belonging to hosts, Rickettsiella strains, and any other microbial organisms infecting the same ticks. For the metagenomes Iric, Ird1, and Ird6, DNA extracted from whole tick bodies was first sequenced using PromethION flow cells (FLO-PRO002) with the Ligation Sequencing Kit (SQK-LSK110) from Oxford Nanopore Technologies, and simultaneously sequenced on the NovaSeq 6000 platform with the Nextera DNA Flex Library Prep Kit (Illumina). For the metagenomes Oerr and Opha, endosymbionts DNA samples, enriched as in (Gottlieb et al. 2015, Duron et al. 2018), were sequenced using HiSeq2000 technology using the TruSeq Nano DNA library construction and HiSeq SBS v3 kits (Illumina).
Genome assembly
Different methods were used for the Ornithodoros and Ixodes samples. For the Oerr and Opha metagenomes, the reads were subjected to a modified version of the Blobology pipeline (Kumar et al. 2013), available on github. The modified version allows an initial filtering of the host reads using the genome of I. scapularis (GCA_016920785.2) as reference, to ease the toll on computing resources, and includes the use of Python3, SPAdes v.3.15.4, bowtie2 v.2.1.0, and blast v.2.12.0. Finally, the resulting genome assemblies were refined with Bandage v.0.8.1 (Wick et al. 2015), by manual removal of contigs unrelated to the assembly graph with no BLAST hits against the R. viridis genome as reference (GCF_003966755.1).
For the metagenomes Iric, Ird1 and Ird6, Minimap2 v.2.24 was used to map all nanopore reads against five known Rickettsiella genome sequences (GCF_000168295.1_ASM16829v1, GCF_000257395.1_ASM25739v1, GCF_001881495.1_ASM188149v1, NZ_AP018005.1 and NZ_CP079094.1). Reads that mapped against the Rickettsiella reference genomes were collected using Samtools v.1.15.1 and then de novo assembled using Flye v.2.9.2_b1786 (error rate 0.03), which resulted in 43 contigs with sizes ranging from 0.5 kb to 1.81 Mb. Medaka v.1.7.2 was used to polish the contigs based on the filtered Rickettsiella Nanopore reads.
The completeness of retrieved Metagenome-Assembled Genomes (MAGs hereafter) was estimated using miComplete v.1.1.1 (Hugoson et al. 2020) (--hmms Bact105). The completeness estimation was performed on a dataset including 12 representative species of Legionellales available on NCBI and using the larger genome of the aquatic Legionellales “Candidatus Berkiella aquae” (GCF_001431295.2) as reference (Supplementary Table 1). This dataset of organisms was used in several analyses in our study and we will therefore refer to it as “Dataset R”. The creation of this dataset was completed on February 1st, 2024. Three R. isopodorum and one R. grylli genome sequences were excluded from the dataset as they do not show significant differences in terms of sequence similarity (ANI higher than 98, calculated with the ANI calculator implemented in EZBioCloud, Yoon et al. 2017) and gene content (no strain specific ortholog clusters) when compared to their reference genomes. A summary of the organisms included in Dataset R and their hosts is presented in Table 2, while more information about the dataset, including the average nucleotide identity among genomes, can be found in Supplementary Table 1.
Table 1 - Collection and processing of samples. * The date 2016 refers to the collection of the ticks in Portugal from which the laboratory colony derived.
|
Tick species |
Strain |
Collection date |
Location |
Environmental context |
Sampling method |
Number of ticks |
Enrichment |
|
Ixodes ricinus |
Iric |
2020 |
Netherlands: De Dorschkamp |
Deciduous forest |
Dragging |
12 (male) |
- |
|
Ird1 |
2021 |
Netherlands: De Amsterdamse Waterleiding Duinen |
Coastal sand dune area |
Dragging |
1 (female) |
- |
|
|
Ird6 |
2021 |
Netherlands: De Amsterdamse Waterleiding Duinen |
Coastal sand dune area |
Dragging |
1 (female) |
- |
|
|
Ornithodoros erraticus |
Oerr |
2016* |
Laboratory colony |
Laboratory colony |
- |
3 (female) |
As described in (Gottlieb et al. 2015, Duron et al. 2018) |
|
Ornithodoros phacochoerus |
Opha |
2021 |
Mozambique: Coutada 9 Game Reserve |
Phacochoerus africanus burrows |
Dragging |
4 (female) |
As described in (Gottlieb et al. 2015, Duron et al. 2018) |
Table 2 - Summary of Dataset R, used for phylogenomics and comparative genomics analyses. The Legionellales representatives’ host scientific and generic names are reported. Strains sequenced in this study are reported in bold.
| Legionellales representatives | Host (if any) | Host type |
| Aquicella lusitana GCF_003350455.1 | Aquatic | Aquatic |
| “Candidatus Berkiella aquae” GCF_001431295.2 | Acanthamoeba polyphaga | Amoeba |
| “Candidatus Berkiella cookevillensis” GCF_001431315.2 | Acanthamoeba polyphaga | Amoeba |
| Coxiella burnetii RSA 493 NC_002971.4 | Dermacentor andersoni | Tick |
| Coxiella endosymbiont of Amblyomma sculptum strain CeAS-UFV CP_033868.1 | Amblyomma sculptum | Tick |
| Coxiella endosymbiont of Ornithodoros maritimus CLEOmar GCA_907164965.1 | Ornithodoros maritimus | Tick |
| Legionella pneumophila GCF_001753085.1 | Homo sapiens | Human |
| Rickettsiella endosymbiont of Dermanyssus gallinae GCF_019285595.1 | Dermanyssus gallinae | Mite |
| Rickettsiella grylli GCF_000168295.1 | Armadillidium sp. | Isopod |
| Rickettsiella isolated from Ixodes ricinus, Ird1 strain | Ixodes ricinus | Tick |
| Rickettsiella isolated from Ixodes ricinus, Ird6 strain | Ixodes ricinus | Tick |
| Rickettsiella isolated from Ixodes ricinus, Iric strain | Ixodes ricinus | Tick |
| Rickettsiella isolated from Ornithodoros erraticus, Oerr strain | Ornithodoros erraticus | Tick |
| Rickettsiella isolated from Ornithodoros phacochoerus, Opha strain | Ornithodoros phacochoerus | Tick |
| “Candidatus Rickettsiella isopodorum” GCF_001881495.1 | Trachelipus rathkii | Isopod |
| Rickettsiella massiliensis 20B GCF_000257395.1 | Ixodes ricinus | Tick |
| “Candidatus Rickettsiella viridis” GCF_003966755.1 | Acyrthosiphon pisum | Aphid |
Genome annotation
Genomes were annotated with Prokka v.1.14.6 (Seemann 2014) using the “compliant” mode and default parameters. The resulting annotations in GenBank format, together with the other 12 annotated Legionellales of Dataset R downloaded from NCBI, were subjected to pseudogene analysis with PseudoFinder v.1.1.0 (Syberg-Olsen et al. 2022) with default parameters. The counts of intact and pseudo-genes have been graphically represented with an in-house R script available on github. KEGG orthology assignments of intact genes and pseudogenes of each genome were predicted using BlastKoala v.3.0 (Kanehisa et al. 2016). The outputs were then merged using in-house scripts available on github. Finally, we screened for mobile elements with MobileElementFinder v.1.1.2 (Johansson et al. 2020) using the database MGEdb v.1.1.1 and default parameters, for prophages with PHASTEST v.3.0 (Wishart et al. 2023), and for defense systems (CRISPR/CAS) with CRISPRCasFinder 1.1.2 (Couvin et al. 2018) as in (Siozios et al. 2024).
Phylogenomics
The phylogenetic relationships between genomes in Dataset R were inferred through a phylogenomic approach on single copy orthologous amino acid sequences (of intact genes, as predicted by PseudoFinder v.1.1.0 (Syberg-Olsen et al. 2022), alignment length 27116 aa) present in all organisms using Methylococcus mesophilus strain 16-5 (GCF_026247885.1, intact genes predicted the same way) as the outgroup. The pipeline is available on GitHub and includes the use of the tools OrthoFinder v.2.5. (Emms & Kelly 2019), SeaView v.5.0.5 (Gouy et al. 2021), ModelTest-NG v0.2.0 (Darriba et al. 2020), RAxML (raxmlHPC-PTHREADS v.8.2.12, Stamatakis 2014, 1000 bootstraps), and in-house scripts in Python3, bash, and R. All of the softwares have been run in default settings, except for RAxML, where the model was set to LG+I+G4 (PROTGAMMAILG), chosen according to the Akaike criterion with ModelTest-NG v0.2.0.
Overall comparative genomics
In order to obtain a description of common and strain-specific genomic features and metabolic capabilities of Rickettsiella, we analysed the clusters of orthologous intact genes detected for Dataset R, regardless of phylogenetic relationships. The OrthoFinder output table from which the single-copy orthologs used in the phylogenomic inference (Orthogroups.GeneCount.tsv, Supplementary Table 2) was analysed for intersections using the R package UpSetR (Conway et al. 2017), and the results were plotted using the same tool. All the commands are available on github.
Particular attention was paid to clusters uniquely shared by (i) all Rickettsiella species and strains, (ii) Rickettsiella strains isolated from hematophagous arthropods, for traces of adaptation to host blood feeding (iii) Rickettsiella strains isolated from phytophagous/detritivorous hosts, for traces of adaptation to other diets (iv) Rickettsiella isolated from the same host, for adaptation to hosts/environments and (v) Rickettsiella and Coxiella isolated from ticks, for traces of adaptation to tick hosts. We also investigated gene orthology in respect to the phylogenetic clusters that appeared well-defined in the phylogenetic trees. Orthology was assessed using OrthoFinder on all translated proteins predicted in genomes by Prokka, as well as on translated sequences of intact genes and pseudogenes as previously predicted with PseudoFinder, in order to obtain more information about the evolution of the strains. The results were graphically represented as Venn diagrams using the R package VennDiagram (Chen & Boutros 2011), and the KEGG and Prokka annotations of the clusters were obtained (commands available on GitHub).
Comparison of functions of interest
We also analysed specific pathways that could be involved in nutritional symbioses with hematophagous arthropods (B vitamins, chorismate and heme metabolism - Zhong et al. 2021, Duron et al. 2018, Hosokawa et al. 2010, Buysse et al. 2021), using subsets of Supplementary Table 3 to obtain graphical representation of the gene presence/absence/pseudogenization along the phylogenomic tree with an in-house R script available on github. The analysis showed differences in some of the pathways. In particular, the thiamine pathway caught our attention: the strain Iric appears to have a set of genes that are absent in other Rickettsiella strains but shared with other Legionellales representatives. We thus further investigated these genes (Supplementary Table 3) by retrieving additional sequences from NCBI, identified via blastp, and aligned the amino acid sequences with MUSCLE v3.8.1551 (Edgar 2004). Finally, we inferred their respective phylogenies with RAxML (100 bootstraps).
We further checked for the presence of virulence factors known in Legionellales (van Schaik et al. 2013, Isberg et al. 2008). Protein sequences of virulence factors annotated in Legionella longbeachae NSW150 (NC_013861), L. pneumophila str. Corby (NC_009494) and L. pneumophila subsp. pneumophila str. Philadelphia 1 (NC_002942) were downloaded from VFDB (Liu et al. 2022) and used as queries in a blastp search against the translated sequences of intact genes and pseudogenes of genomes from Dataset R.
Finally, we also looked for the presence of CI genes. The CI phenotype is encoded by a pair of syntenic genes, cifA and cifB, whose direct genotype-phenotype association has been demonstrated in Wolbachia (LePage et al. 2017, Beckmann et al. 2019). cifA and cifB possess homologs that have been categorised into five phylogenetic clusters (Martinez et al. 2021, Martinez et al. 2022, Vancaester & Blaxter 2023, Tan et al. 2024), although type V has recently been split into type V-to-X (Tan et al. 2024). To determine the presence of cif gene homologs in the Rickettsiella genomes, we used Orthofinder to search for specific orthologs of the cif gene groups identified in a previous study (Martinez et al. 2021). These searches were conducted on a database of CDSs from Dataset R, as well as from published Wolbachia, known to induce CI, and Rickettsia genomes (Supplementary Table 4). Protein domains were predicted for the cifA and cifB homologs detected using the HHpred webserver (https://toolkit.tuebingen.mpg.de/#/tools/hhpred/, Söding et al. 2005) with default parameters (Lindsey et al. 2019), and using the following databases: SCOPe70 v.2.08, Pfam v.36.0, SMART v.6.0, and COG/KOG v.1.0. The sequences of each detected domain were retrieved and manually aligned using Clustal Omega (Sievers & Higgins 2018), independently for each cifA and cifB homologs, to complete the domain detection of some false negatives generated by HHpred. Visualisation of protein domains within the cif genes was performed using RStudio v4.3.1 with the ggplot2 v.3.4.4 (Wickham 2009), cowplot v1.1.1 (Wilke 2024), and gridExtra v.2.3 (Auguie 2017) packages. A maximum likelihood phylogeny of cifA and cifB pair genes has been inferred on the basis of three conserved domains (RNA-binding-like, AAA-ATPases-like and PD-(D/E)XK nucleases) independently aligned and concatenated. The phylogenetic reconstruction was performed using the best-fitting substitution model, here JTT+G, as identified by ModelTest-NG, based on the Akaike Information Criterion.
Results
Rickettsiella metagenomes
We sequenced and assembled three microbial metagenomes from I. ricinus (Ird1, Ird6 and Iric), one from O. phacochoerus (Opha) and one from O. erraticus (Oerr). We obtained an average of 71.86 Gb of paired-end reads (from 2.2 gb to 192 bp) of 150 nt in length for each sample (see Supplementary Table 1 for details). For each metagenome, we extracted the reads belonging to the target organisms and obtained the sequences of five new Rickettsiella MAGs (see Supplementary Table 1 for details). They are all of similar size (~1.7 Mb), with the exception of Rickettsiella Oerr MAG (~1.3 Mb), and most of them are estimated to be complete. However, the estimation attributes to Rickettsiella strain Oerr MAG lower completeness scores (0.9048; weighted completeness 0.8109) among the MAGs sequenced in this study. Lower scores are expected for endosymbiotic, host-restricted bacteria, whose genomes undergo erosion (Moran 2002, McCutcheon & Moran 2011, Boscaro et al. 2017, Kashkouli et al. 2021), and observed in Rickettsiella relatives, as the Coxiella-like endosymbionts of ticks. Thus, the completeness score of Rickettsiella strain Oerr MAG reflects more likely genomic erosion than poor assembly, albeit the number of contigs included in this MAG reaches 99. However, it is worth noting that other genomes, previously published and considered to be complete, are assembled in a higher/comparable number of contigs (Coxiella CLEOmar, Aquicella lusitana) or present a lower set of markers (Coxiella CeAS-UFV, 93 out of 105 markers used in the analysis, two less than Rickettsiella Oerr). The GC content varies between 36% and 39%, values also consistent with those from endosymbiotic bacteria. Metagenomes also contain reads of other bacterial endosymbionts as Midichloria and Francisella-like endosymbiont for I. ricinus and O. phacochoerus, respectively (data not shown). These endosymbionts were not further considered in this study. No other endosymbiont than Rickettsiella was detected for O. erraticus.
The annotation of the five Rickettsiella MAGs revealed a somewhat similar number of functional genes, ranging from 1,268 to 1,473 (Supplementary Table 1). Including all Rickettsiella genomes shows that the number of functional genes is between 1,189 for R. isopodorum and 1,707 for the Rickettsiella endosymbiont of D. gallinae (Supplementary Table 1, Supplementary Figure 1). Rickettsiella genomes display similar levels of pseudogenization to other Legionellales, though some variation is observed. For instance, higher numbers of pseudogenes were predicted for the Rickettsiella MAGs Iric and Oerr (340 and 237 respectively), R. massiliensis 20B (508) and a few other Legionellales, including Coxiella-like endosymbiont of Ornithodoros maritimus strain CLEOmar (778). In contrast, lower numbers of pseudogenes were predicted in other Legionellales, such as the Coxiella-like endosymbiont of Amblyomma sculptum strain CeAS-UFV (Leclerque et al. 2012) (Supplementary Table 1, Supplementary Figure 1). Concerning mobile elements, only one was detected in the analysed dataset, and only in R. grylli (Supplementary Table 1). Both recently host-restricted bacteria and long-term symbionts generally possess a low number of mobile elements in their genomes, although recent symbionts are usually characterised by a greater number. However, long term symbionts are additionally characterised by fewer pseudogenes and smaller genome sizes (McCutcheon & Moran 2011) than the Rickettsiella strains here investigated. Taken together, these features suggest a relatively recent infection of the hosts by Rickettsiella. In addition to mobile elements, prophage sequences were detected in the strains Iric and Ird6 and in the Rickettsiella endosymbiont of D. gallinae (Supplementary Table 1), but not in other strains. Furthermore, we investigated defense systems to evaluate the degree of adaptation to an endosymbiotic lifestyle as in Arsenophonus (Siozios et al. 2024) (Supplementary Table 1).
Most Average Nucleotide Identities (ANIs) between the five Rickettsiella MAGs range from 69.39 to 72.61%, suggesting that they may be distinct species (Supplementary Table 1). A notable exception is the 99.77% ANI between Iric and Ird6. The five Rickettsiella MAGs have ANIs ranging from 69.36-to-98.46% with other Rickettsiella genomes. Indeed, ANI is 98.46% between Iric/Ird6 and R. massiliensis, and 98.26% between Ird1 and R. isopodorum (Supplementary Table 1).
Phylogenomics of Rickettsiella
Phylogenomic analysis based on 109 single-copy orthologs showed that Rickettsiella, Coxiella, Legionella, Aquicella, and Berkiella genera cluster together in a robust monophyletic clade forming the Legionellales order (Figure 1). The inner tree topology is also congruent with the known genera of Legionellales (Duron et al. 2018). Indeed, genomes of Rickettsiella, Coxiella, and Berkiella cluster together according to their assigned genus and form robust monophyletic subclades, with Aquicella and Coxiella forming sister genera of Rickettsiella. Two Rickettsiella MAGs (Iric and Ird6) obtained in this study from I. ricinus, cluster with R. massiliensis sharing a high ANI from 98.5 to 99.8% (node 15, Figure 1, and Supplementary Table 1), suggesting that these three Rickettsiella belong to the same species. The three other Rickettsiella MAGs obtained in this study cluster with diverse Rickettsiella species: Opha with R. viridis, the defensive symbiont of aphid, and the putative nutritional Rickettsiella symbiont of poultry red mite (node 13), Ird1 with R. isopodorum (node 12) and Oerr with a group formed by R. grylli, R. isopodorum and Irid6 (node 10, Figure 1). The Rickettsiella found in different tick species are thus distantly related and do not form a monophyletic group (Figure 1).
Comparative genomics
Orthology analysis
The analysis of the intact orthologous genes detected for Dataset R (Figure 2, Supplementary Figure 2, Supplementary Table 2) indicates huge variation among Legionellales, with only135 clusters shared by all, most of which are involved in basic bacterial processes. Only 25 clusters of intact genes are shared exclusively by members of the Rickettsiella genus, most of which are functionally uncharacterised (Supplementary Table 2). No clusters of intact genes specific to Rickettsiella and Coxiella strains isolated from hematophagous arthropod hosts were detected. The core Rickettsiella genome appears to contain 606 COGs, that drop to 362 when pseudogenes are removed from the analysis; pseudogenes in the Rickettsiella core genome (29 clusters of pseudogenized sequences) appear to be mainly associated with transcription and regulation functions (Supplementary Table 5). We then attempted to detect gene families specific to the phylogenetic groups that could be identified on the phylogenomic tree shown in Figure 1.
Node 10 encompasses two previously described and potentially pathogenic Rickettsiella species isolated from isopods (R. grylli - Leclerque 2008 - and R. isopodorum - Kleespies et al. 2014), and two of the newly Rickettsiella strains identified in ticks. Notably, these two new strains were isolated from very distant ticks, both in terms of host species and geographic location (Supplementary Table 1). The core genome of these organisms contains 823 COGs, which drop to 704 when pseudogenes are not considered in the analysis (Figure 1). Of the 704 COGs, 198 correspond to uncharacterised proteins, and 27 appear to be involved in secretion of effectors and pumps. Furthermore, five were annotated by Prokka as multidrug resistance proteins. However, the putative pathogen R. grylli possesses 22 additional transport proteins compared to the rest of this dataset, and Rickettsiella Oerr shows 35 specific uncharacterised proteins (Supplementary Table 5).
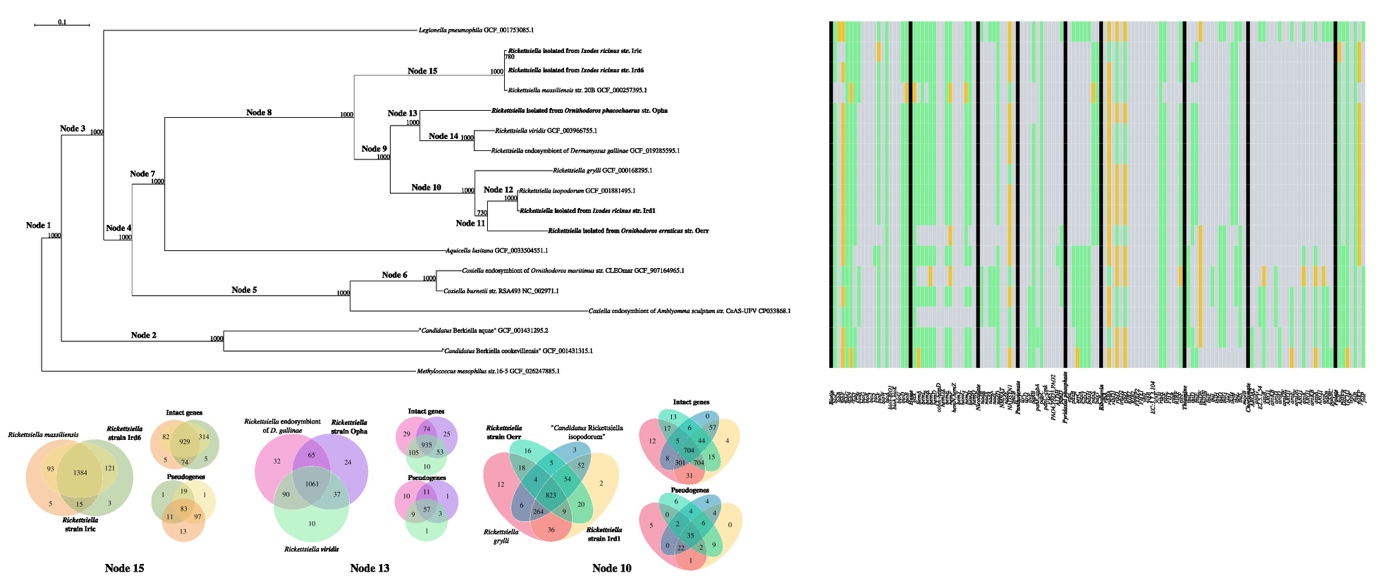
Figure 1 - Phylogeny of Legionellales (Dataset R) focusing on the Rickettsiella genus inferred with RAxML on 109 single copy orthologous intact genes present in all organisms in the dataset (1000 bootstraps, evolutionary model: LG+I+G4, rapid bootstrap random number seed: 1234, the random number seed for the parsimony inferences: 123), using Methylococcus mesophilus as outgroup to root the tree. The Rickettsiella organisms whose genomes were sequenced in this study are indicated in bold. On the right, the presence (green), absence (grey) and pseudogenization (yellow) of genes of specific metabolic pathways annotated with KEGG is represented as a heatmap. M. mesophilus sequences were not annotated. On the bottom, Venn diagrams of clusters of orthologs of all encoded genes, intact genes, and pseudogenes is provided for the specified nodes of the tree (Node 10, Node 13, Node 15).
Node 13 encompasses the host colour-altering and defensive symbiont R. viridis (aphid host, Tsuchida et al. 2014), the Rickettsiella endosymbiont of D. gallinae (poultry red mite host, De Luna et al. 2009), and the newly sequenced Rickettsiella str. Oerr isolated from the soft tick O. phacochoerus. The core genome of this node contains 935 COGs (1061 when pseudogenes are considered; Figure 1). All the Rickettsiella strains in this node appear to maintain a discrete, albeit low, number of unique COGs. Among others, the core intact genes include T4SS components, ATPase subunits, biotin genes, 79 hypothetical proteins, four ankyrin repeat domain-containing proteins, and 14 DUF domain-containing proteins. Notably, the Opha strain appears to share more COGs with the Rickettsiella of D. gallinae (65, encoding hypothetical proteins, several ATPases, DUF containing-proteins, and transcription regulators) than with R.viridis (37, most of them being potentially involved in adhesion). Interestingly, the core pseudogenes include many response regulators and translocators. Finally, the aerobic respiration control sensor protein ArcB is pseudogenized in the strain Opha only, and the endosymbiont of D. gallinae contains 10 more unique pseudogenes, that include regulators and transposases (Supplementary Table 5).
Finally, Node 15 encompasses three Rickettsiella strains isolated from the hard tick I. ricinus: R. massiliensis, Rickettsiella Iric and Ird6. The core of 929 COGs (1384 when pseudogenes are included in the analysis) contains mainly hypothetical proteins and house-keeping genes, but also genes involved in energy production. Interestingly, the strain Iric exhibits distinctive COGs exclusively when pseudogenes are analysed, revealing a single cluster associated with amino sugar and nucleotide sugar metabolism (Supplementary Table 5). However, the other strains possess only five unique COGs as well. A relatively high number of intact sequences is shared by any two organisms in these subsets of genomes. Most sequences shared by R. massiliensis and the strain Iric are annotated as hypothetical proteins, whereas most sequences shared by R. massiliensis and the strain Ird6 are housekeeping genes and genes encoding chaperones. The Rickettsiella Iric and Ird6 share mostly uncharacterised proteins, but also several dehydrogenases, ATPase synthase subunits A and B, and a queuosine precursor transporter (Supplementary Table 5).
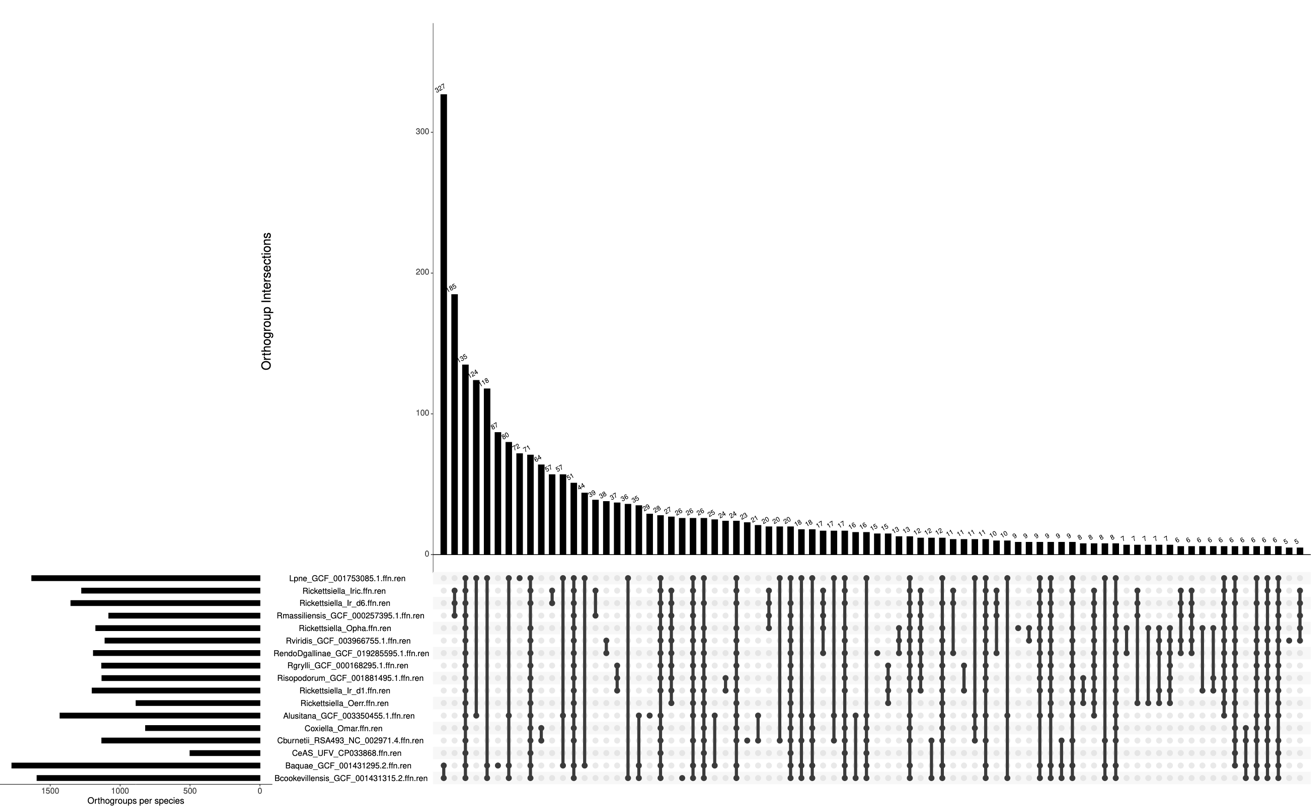
Figure 2 - Intersection plot displaying shared and specific clusters of orthologous proteins between Rickettsiella and other Legionellales. Intersection plot is based on Dataset R and was produced with the R package UpSetR. In the left side, the horizontal bar plot represents the number of orthogroups present in each organism of the dataset. In the top, the vertical bar plot represents the number of orthogroups that belong to specific intersections of organisms in the dataset. The height of each bar indicates how many orthogroups are in that particular intersection, i.e., shared by the combination of organisms indicated by the intersection matrix below. In the intersection matrix, on the bottom, each row represents an organism and each column represents a possible intersection of these organisms. Filled circles indicate that the organisms in that row are part of the intersection shown in the corresponding column. The lines connecting the filled circles indicate which organisms are included in each specific intersection. This plot is limited to six orthogroups in the intersections, for the full plot of all intersections until one shared orthogroup please see Supplementary Figure 2.
Metabolic capabilities
The annotation of metabolic pathways (Supplementary Table 3) shows overall similar metabolic capabilities among the members of the Rickettsiella genus. Major differences can be identified, however, in specific pathways: the respiration and ATP production appears to be most degraded in strain Iric and R. massiliensis 20B, i.e., the two Rickettsiella strains with the higher number of predicted pseudogenes that form a monophyletic cluster together with the Ird6 strain. Furthermore, B vitamins and heme pathways show significant differences, appearing most degraded in R. massiliensis 20B, but still relatively conserved in the strain Iric (e.g., biotin, folate). Strikingly, the heme biosynthesis pathway is absent in the strain Oerr, which has also lost most of the thiamine biosynthesis pathway, while retaining a unique segment of this pathway that is not found in other Rickettsiella strains sequenced in this study. However, although it is not possible to fully reconstruct the evolution of these sequences, a blast search and inferred phylogenies for each gene shows that they are of Gammaproteobacteria origin (Supplementary Table 3, five best hits shown, and Supplementary Figure 3), often found in Legionella spp. and in two additional Rickettsiella organisms. This suggests either multiple horizontal transfers from other Gammaproteobacteria or Legionellales, or a loss of the genes in the strains analysed here. Finally, while Coxiella-like endosymbionts can produce chorismate, a tryptophan precursor that regulates serotonin biosynthesis in ticks (Ríhová et al. 2017), Rickettsiella genomes do not harbour genes for this biosynthetic pathway, suggesting that they have no impact on tick behaviour through this mechanism.
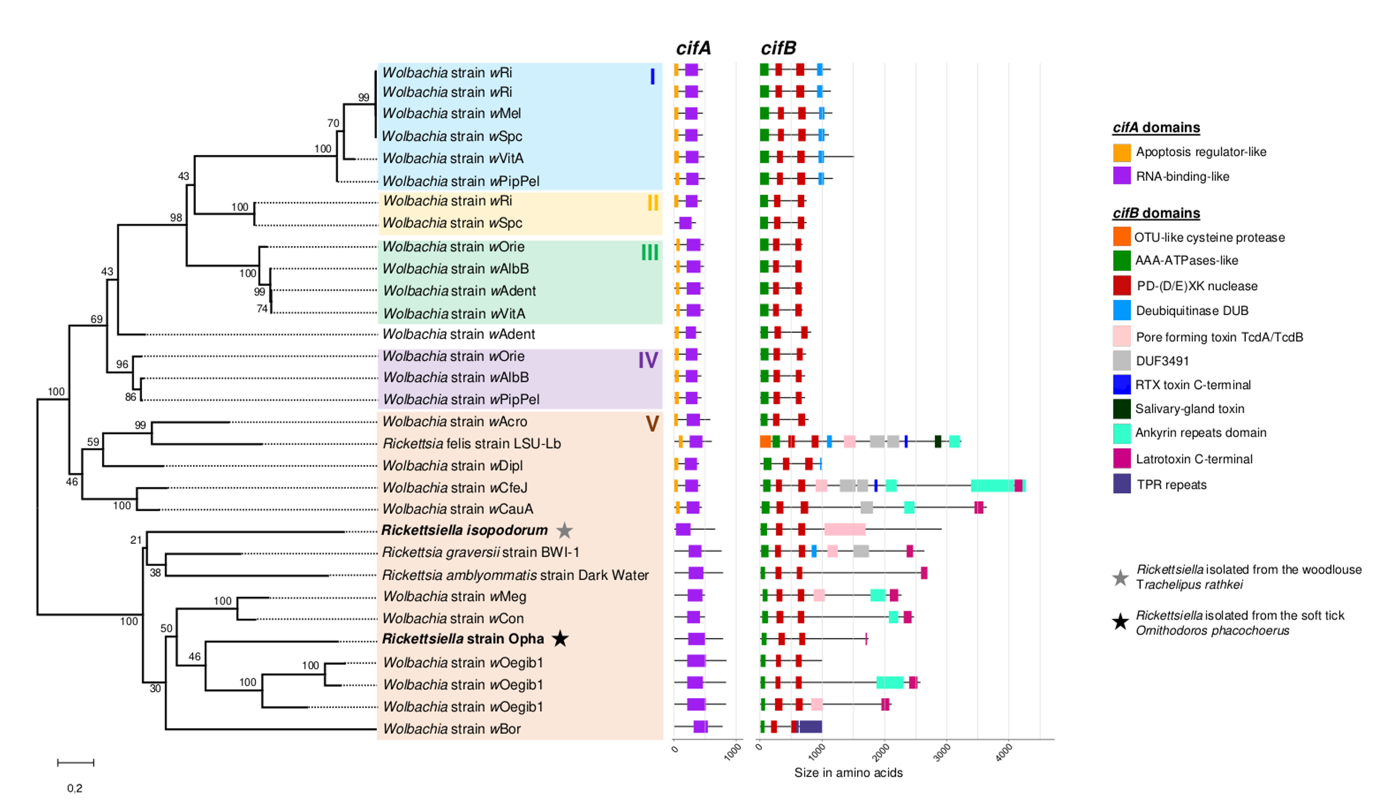
Figure 3 - Phylogeny and domain architecture composition of cifA and cifB homologs in Rickettsiella and other endosymbionts. The maximum-likelihood tree was reconstructed from an alignment concatenated with RNA-binding-like, AAA-ATPase-like, and PD-(D/E)XK nuclease domain sequences (191 amino acid positions, JTT+G substitution model). The tree is midpoint rooted. The type I-V cif representation, as well as the sizes and domain compositions of the cifA and cifB genes, were drawn relative to those described in Martinez et al. (2021, 2022). The scale bar represents the inferred number of substitutions per site. Bootstrap values were estimated from 1000 replicates. Accession numbers of the Rickettsiella, Wolbachia, and Rickettsia genomes used in this analysis are listed in Supplementary Table S4.
Identification of virulence factors
Further genomic comparisons showed that abundance of virulence genes varies between Legionellales members (Figure 4, Supplementary Table 6). While large repertories of virulence genes associated with adherence, secretion systems, effectors and exotoxin RtxA are common in L. pneumophila genomes, these genes are rare or absent in most other Legionellales as Coxiella-like endosymbionts associated with ticks. Indeed, some virulence genes are absent in Rickettsiella genomes including the exotoxin RtxA-encoding locus, which is involved in adherence, cytotoxicity, and pore formation in vertebrate cells by Legionella species (Cirillo et al. 2001). Aquicella lusitana also lacks the exotoxin, suggesting that the loss of this gene might be ancestral to Rickettsiella and Aquicella. As with the other factors associated with virulence in Legionella, Rickettsiella genomes conserve those involved in adherence, regulation, and nutritional metabolism (particularly, heme metabolism cofactors, Shevket et al. 2018). Furthermore, the Rickettsiella strains retain the effector delivery systems, but not the secreted virulence-associated effectors identified in Legionella, suggesting that this machinery could be exploited by Rickettsiella to perform different functions. The same applies to Aquicella, Berkiella and C. burnetii, whereas the two Coxiella-like endosymbionts have neither the delivery system nor the genes encoding for effectors. Furthermore, all the investigated Legionellales, with the exception of Legionella and several Rickettsiella strains, do not possess the Type IV A secretion system (TIVASS). The strains that maintain most of this system are R. viridis and the tick strains Ird1, Oerr and Opha, while the Rickettsiella endosymbiont of D. gallinae, R. grylli and R. isopodorum have only a few genes involved in this machinery. No evidence of horizontal gene transfer was found. Finally, no enhancer-binding protein (Enh) encoding loci were found in Rickettsiella, whereas they are found in Aquicella, Berkiella, C. burnetii, and Legionella.
Rickettsiella and cytoplasmic incompatibility
We detected homologs of the CI genes cifA and cifB in the genomes of (i) Rickettsiella strain Opha, and (ii) R. isopodorum where a CifB-like protein has been previously identified (Figure 3, Gillespie et al. 2018). Both strains harbour the RNA-binding-like domain in cifA, and the AAA-ATPase-like and PD-(D/E)XK nuclease domains in cifB, both conserved in all known cif genes (Martinez et al. 2021, Beliavskaia et al. 2023). Additionally, the cifB gene in the strain Opha has a weakly aligned latrotoxin-like fragment, while R. isopodorum has a large TcdA/TcdB pore-forming toxin domain, also found in other cifB homologs of Wolbachia and Rickettsia. Furthermore, we also identified a unique cifB homolog (without cifA) in a Rickettsiella sp. genome reconstructed from eDNA isolated from hot springs and volcanic lakes (GCA_037439325.1) and in the distant relative “Ca. Aquirickettsiella gammari” (GCA_002290645.2).
Both the detected cifA and cifB homologs of Rickettsiella cluster within the Type V cif paraphyletic group, which is the most diverse in terms of size and domain composition (Martinez et al. 2021, Beliavskaia et al. 2023). These genes are located in the chromosomal DNA and without any nearby mobile genetic elements, such as prophages, insertion sequences (IS), or retrotransposons. Both the cif genes from Rickettsiella strain Opha and R. isopodorum group together in a highly supported clade comprising cif genes from Rickettsia and Wolbachia, originating from at least two supergroups (wOegib and wBor from supergroup A, wMeg and wCon from supergroup B), showing the occurrence of gene transfers between these phylogenetically distinct endosymbionts. However, the long branch lengths and low intra-group bootstrap values create uncertainty regarding the exact position of the Rickettsiella cif genes, challenging the reconstruction of their evolutionary history and origin.
Discussion
We show here that members of the genus Rickettsiella share a set of similar metabolic capabilities, as pathways involved in energy production. Most notably, the Rickettsiella strains lack most key virulence genes responsible for persistence and replication in vertebrate cells of pathogenic Legionella and Coxiella. Indeed, while the Rickettsiella genomes harbour the virulence genes encoding the Dot/Icm secretion system, the genes encoding the Legionellales Dot/Icm secreted effectors and the exotoxin RtxA have been pseudogenized or are completely absent. However, examination of Rickettsiella genomes further reveals a greater variability in metabolic properties associated with endosymbiosis: They have variable capacity to biosynthesize de novo certain B vitamins (biotin, riboflavin, pyridoxal phosphate, pantothenate) and heme, and some harbour CI genes, suggesting that they are engaged in complex interactions with their respective arthropod hosts. Phylogenomics further confirmed that members of the genus Rickettsiella form a robust monophyletic clade of Legionellales associated with arthropods but that tick-borne Rickettsiella do not cluster within a specific subclade. Tick-borne Rickettsiella rather exhibit distinct evolutionary origins, revealing that Rickettsiella have undergone repeated horizontal transfers between ticks and other arthropods, including aphids and woodlice.
The virulence factors presence/absence pattern across Legionellales reveals key past events in the evolutionary history of Rickettsiella. Indeed, the absence of major Legionellales virulence factors in all Rickettsiella strains, including the putative human pathogen R. massiliensis 20B, suggests that these microorganisms are not strongly pathogenic to mammals as L. pneumophila and C. burnetii. Aquicella lusitana, Rickettsiella’s closest relative, also lacks the RtxA exotoxin, suggesting that the loss of this gene could be ancestral to Rickettsiella and Aquicella and extend deep into the evolutionary past of the Coxiellaceae. Several Legionella virulence factors are conserved in Rickettsiella and to some extent in Aquicella, though they may have alternative functions. Notably, traits such as adherence, stress response regulation, nutritional metabolism (particularly involving heme cofactors), and the type IV secretion systems are common to both pathogenic and mutualistic bacteria (Kashkouli et al. 2021, Cirillo et al. 2001, Shevket et al. 2018, Gillespie et al. 2018, Beliavskaia et al. 2023, Lomovatskaya & Romanenko 2020, Costa et al. 2021, Cao et al. 2018, Floriano et al. 2023) whereas the Dot/Icm effector delivery system has been so far exclusively associated with pathogenesis (Santos-Garcia et al. 2023). Indeed, genes encoding the Dot/Icm system are either absent or pseudogenized in Coxiella-like endosymbionts associated with nutritional symbiosis in ticks (Santos-Garcia et al. 2023, Buysse & Duron 2021). The conservation of this secretion system in Rickettsiella suggests either that this machinery is exploited by Rickettsiella to fulfill functions other than virulence by secreting different effectors, or that Rickettsiella maintains a degree of virulence but uses different virulence effectors to those of Legionella. Alternatively, since the Rickettsiella Dot/Icm system lacks a few genes compared to L. pneumophila and C. burnetii, this machinery may have undergone a gradual degradation and they mirrored a global genome decay, as commonly observed in endosymbionts of arthropods (Moran 2002, McCutcheon & Moran 2011, Kashkouli et al. 2021, Klasson & Andersson 2004, Bennett & Moran 2015). Whatever the scenario, virulence against mammals seems to be gradually lost in Rickettsiella, leaving opportunities for the evolution of other interactions with their arthropod hosts.
The genomes of Rickettsiella consistently harbour genes involved in narrow interactions with arthropods, although these genes vary between trains and species. The biosynthesis genes of several B vitamins (biotin, riboflavin, and folate) were found intact in most Rickettsiella genomes, suggesting that they could supply these nutrients to their arthropod hosts. In this context, Rickettsiella may be essential mutualistic partners for hematophagous arthropods, such as ticks and certain mites, which need these essential vitamins for their growth and the completion of their life cycle (Duron & Gottlieb 2020). Most tick species harbour Coxiella-like endosymbionts which provide B vitamins, but tick species lacking Coxiella-LE instead host Francisella-like endosymbionts (Duron & Gottlieb 2020, Binetruy et al. 2020b) or Midichloria (Duron 2024) as we observed in this study in O. phacocherus and I. ricinus, respectively. These symbionts are also capable of supplying B vitamins to their hosts. Co-infections (i.e., the simultaneous infection of Rickettsiella with one or more different microorganisms) have been previously documented in ticks (Duron et al. 2017, Duron et al. 2015a, Binetruy et al. 2020b), hence, the identification of Rickettsiella tick species previously associated with other symbionts is not surprising. However, in O. erraticus, we did not observe any other endosymbionts than Rickettsiella which suggests that this bacterium is an obligate nutritional partner of this host. Current knowledge, including a study on a related tick species (Duron et al. 2018), shows that a mutualist is necessary. In addition, certain Rickettsiella genomes possess biosynthesis genes for heme, a nutrient which is essential for regulating protein synthesis and cell differentiation in animals. Unlike other eukaryotes, ticks lack most of the genes encoding proteins necessary for heme production and degradation (Jia et al. 2020, Gulia-Nuss et al. 2016, Hatalová et al. 2023). Some Rickettsiella endosymbionts could thus potentially complement tick metabolism through the production of this essential cofactor.
Further examinations of Rickettsiella genomes also suggest that at least two strains may be reproductive parasites of the soft tick O. phacochoerus and the woodlouse Trachelipus rathkei. Although no phenotypic data are currently available for these arthropod species, their Rickettsiella possess CI-inducing genes with key functional domains, including a RNA-binding-like domain in cifA, a the AAA-ATPase-like and PD-(D/E)XK nuclease domains in cifB, as observed in all cif genes of CI-inducing Wolbachia (Martinez et al. 2021, Beliavskaia et al. 2023). The observed pattern suggests acquisitions of cif genes in Rickettsiella through lateral gene transfers from phylogenetically distant endosymbionts also inhabiting arthropods, like Rickettsia and Wolbachia. Both are common in arthropods (Weinert et al. 2015, Duron et al. 2008), especially Rickettsia in ticks (Duron et al. 2017, Binetruy et al. 2020a) in which co-infections with Rickettsiella have been reported (Duron et al. 2017). These co-infections create physical proximity between these endosymbionts and may have facilitated lateral DNA transfers (Bordenstein & Wernegreen 2004, Duron 2013), which could explain the intertwined evolutionary history of cifA and cifB genes among endosymbionts of arthropods.
The phylogenetic diversity pattern strongly suggests that horizontal transfers between arthropod species are key drivers of Rickettsiella spread, dictating global incidence of infections across arthropod communities. Repeated horizontal transfers results to a widespread and global distribution in diverse arthropod hosts, but also to multiple acquisitions of different Rickettsiella strains in the same host species as in I. ricinus (this study) and the polar seabird tick Ixodes uriae (Duron et al. 2016). The ability of Rickettsiella to switch from one host species to another may therefore explain why no COGs specific to tick-borne Rickettsiella have been identified. There are, however, some exceptions: i) one uncharacterised protein is shared by all Rickettsiella isolated from I. ricinus, ii) the fatty acid oxidation complex subunit alpha is shared by all strains isolated from I. ricinus apart from R. massiliensis 20 B, iii) five clusters of hypothetical proteins specific to the Oerr strain, and nine clusters of uncharacterised proteins specific to the Opha strain. While most of these proteins have not been associated with any function yet, the fatty acid oxidation complex is quite interesting. Indeed, bacteria may exploit fatty acids from the host and use them for their own sustainment (Sonenshine & Roe 2013). On the other hand, the oxidation of fatty acids might allow Rickettsiella to produce energy-rich compounds that could be used by Rickettsiella itself but also its tick host, aiding the host’s metabolism, as previously suggested for the tick microbiome (Narasimhan & Fikrig 2015, Board on Population Health 2011) and Coxiella-like endosymbionts (Gottlieb et al. 2015, Bonnet et al. 2017).
Conclusions
Rickettsiella is a diverse group of endosymbiotic bacteria that exhibit a range of adaptations, from pathogenic to mutualistic, enabling them to thrive within arthropod cells and manipulate their hosts’ biology. Over the past decade, significant progress has been made in understanding their impact on arthropods and introducing these bacteria into the field of genomics. Our study highlights that Rickettsiella strains share similar metabolic capabilities and collectively lack virulence genes, suggesting a functional role in the arthropod hosts. In addition, we identified homologs of cif genes, associated with cytoplasmic incompatibility in Wolbachia, in two Rickettsiella strains, suggesting that they may use reproductive manipulation to spread in host populations (as previously shown for another Rickettsiella strain isolated from spiders). However, Rickettsiella remains an understudied aspect of tick biology, with tick-associated strains not forming a monophyletic clade, and a comprehensive approach is needed to fully understand its interactions with various arthropod hosts. Key questions remain, such as: How are the abundance, diversity, and distribution of Rickettsiella maintained worldwide? Can Rickettsiella be used effectively in pest control? How do they manipulate host reproduction? What roles do they play in the evolution of their arthropod hosts? Importantly, future research is needed to further investigate how Rickettsiella affects the fitness of their arthropod hosts, including ticks, and to what extent they can cause diseases in humans.
List of abbreviations
|
Ca. |
Candidatus |
|
CI |
Cytoplasmic incompatibility |
|
R. |
Rickettsiella |
|
TCA |
Tricarboxylic acid (cycle) |
|
TIVASS |
Type IV A secretion system |
|
min |
minutes |
|
nt |
Nucleotides |
|
ANI |
Average Nucleotide Identity |
|
CDSs |
Coding sequences |
|
COGs |
Clusters of Orthologous Genes |
|
aa |
Amino acids |
Acknowledgements
We thank Carlos João Quembo, Florian Taraveau and Maxime Duhayon for their contribution in the sample collection. This work was performed using the computing facilities of the CC LBBE/PRABI. Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Evolutionary Biology: https://doi.org/10.24072/pci.evolbiol.100853 (Jousselin 2025).
Data, scripts, code, and supplementary information availability
The genomes are available on ENA under project number PRJEB70514 (Ird1 and Ird6) and PRJEB82506 (Iric, Oerr and Opha).
All the bioinformatics pipelines and supplementary information are available (https://github.com/annamariafloriano/RickettsiellaComparative, https://doi.org/10.5281/zenodo.17251065 (Floriano 2025).
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article. The authors have complied with all ethical standards required for conducting this research. Consents and approvals are not applicable to this research.
Funding
This work was funded by French Agence Nationale de la Recherche (ANR, France, ref. ANR‐21‐CE02‐0002, Laboratoire d’Excellence CEBA, ref. ANR‐10‐LABX‐25‐01 and LabEx CeMEB, ref. ANR‐10‐LABX‐04‐01), and by the United States Department of Agriculture (USDA, USA, grant no. 2019-67015-28981; https://www.asf-nifnaf.org/).