
 CC-BY 4.0
CC-BY 4.0
Introduction
Processes behind adaptation and speciation have long been limited to recombination and mutation. Only during the last decades, hybridisation has been increasingly acknowledged as another significant driver of evolution, initially mostly in plants but now there is no doubt that hybridisation is of general importance also for animal species diversity (Mallet, 2007; Abbott et al., 2013). Hybridisation and its effects on speciation in animals is often less clear cut than in plants, as much of hybridisation involves successive introgression of genes, in large part due to the general lack of polyploidy in most animal taxa (Mallet, 2005; Schumer et al., 2014). If isolation mechanisms are overcome, genetic variation of the resulting hybrids can lead to the development of novel traits, which differentiate the hybrids from their parent species or parent populations (Mallet, 2007). These novel traits may preadapt hybrid populations to occupy novel niches which e.g. arise due to climate change or anthropogenic influences (Seehausen, 2004).
Hybrids not only need to cope with the physical but also with the biological environment e.g. through coping with resource competition against existing species. Success in competition is frequently observed and the result of heterosis or hybrid vigour, which describes the increased fitness that first generation hybrids may experience compared to their parents, due to favourable recombination of parental traits. Hybrid vigour would not arise if traits were instead exhibited as an addition of those of the parents (Birchler et al., 2010). On the other hand, the new genetic combination may also lead to a decrease in fitness in the form of outbreeding depression (Edmands, 2007), which might cause inviable or infertile hybrid offspring. Predicting whether hybrid offspring will exhibit hybrid vigour or outbreeding depression is difficult without large scale breeding and testing of the hybrids (Orr & Turelli, 2001).
In cases where hybridisation leads to a novel genetic subpopulation, geographical range expansion of the species as a whole can be a result (Pfennig et al., 2016). Due to various phenotypic or genetic population differences and the resulting variation of isolation mechanisms, it is possible for directed hybridisation to occur (Pfennig, 2021). Under directed hybridisation, specific combinations of parents will produce the most fit offspring, which will therefore form the main stock of a hybrid population. These processes can be heavily influenced by sexually selected traits in the parents (Boughman et al., 2024; Franke & Schneider, 2024).
A recent incidence of hybridisation with accompanying range expansion occurred in the wasp spider Argiope bruennichi. Likely due to climatic changes, Asian and Mediterranean European populations of A. bruennichi met and hybridised along a hybrid zone in Central and Eastern Europe (Krehenwinkel & Tautz, 2013; Krehenwinkel et al., 2016) and the species has expanded rapidly in the last half century across Central and Northern Europe (Kumschick et al., 2011).
The resulting Northern European population exhibits several adaptations allowing the spiders to cope with the different climate in the novel habitat. Genetic analyses have exposed the introgression of Asian alleles into a base population of Southern European spiders as responsible for the cold tolerance of the new population (Krehenwinkel et al., 2015; Sheffer et al., 2024). A previous study showed the importance of A. bruennichi’s complex mating system on the likely direction of the hybridisation process and could rule out any prezygotic isolation mechanisms, while suggesting European females and Asian males as the most successfully hybridising partners, and thus the ancestors of the new northern population (Franke & Schneider, 2024). While it is clear that introgression occurred from Asian spiders, the geographical origin of these remains unknown, although Central Asia with its harsh winters has been suggested as likely (Krehenwinkel et al., 2015).
There are several environmental factors that differentiate the new northern habitat from formerly occupied ones, most of which required adaptations in A. bruennichi to facilitate successful colonisation. A. bruennichi is a seasonal spider, overwintering in densely woven egg sacs, emerging from these egg sacs in spring, and maturing and reproducing during summer (Guttmann, 1979). While in their egg sacs, the spiderlings are subject to the natural temperature fluctuations that occur in European and Asian winters. In addition to being insulated against cold temperatures by the egg sac’s silk, spiderlings produce various metabolites that act as cryoprotectants, preventing freezing during temperatures below 0°C. Physiological analyses of the freezing points of spiderlings collected from different regions of the native and expanded habitat showed pronounced differences in their concentrations, resulting in more frost-tolerant spiders in the northern regions (Ortiz-Movliav et al., 2024; Sheffer et al., 2024). Meanwhile, spiders from the more southern regions had lower concentrations of these metabolites, such as fatty acids, amino acids, and cyclic polyols, and consequently showed higher freezing points.
While the environmental cues that lead to spiderlings emerging from the egg sacs are not definitely known for A. bruennichi, both increases in day length and temperature during spring play an important role in the development of other temperate arthropod taxa (Tauber et al., 1986; Roy et al., 2002; Saunders et al., 2002) and are likely to influence A. bruennichi too. Timing the emergence correctly is vital for the spiderlings, as emerging too early may lead to death due to late frosts, while emerging late has the potential disadvantage of missing out on abundant prey of the right size, as well as maturing later than competitors when it comes to finding mating partners (Zimmer et al., 2012).
Due to the differences in summer day length and temperature between Southern and Northern European habitats, A. bruennichi has developed adaptive differences in timing their sexual maturation between the populations (Cory & Schneider, 2018b). While southern spiders are able to take advantage of a long warm period with abundant prey, northern spiders are more time restrained. This has led to northern spiders adapting a more synchronized maturation, with females reaching sexual maturity early in the mating season. Meanwhile, female A. bruennichi of Southern Europe mature less synchronized and freshly matured virgin females are available to males for longer, extending the mating season. Males of both populations benefit from early maturation, and are therefore protandrous and appear early in the season in all populations (Fromhage et al., 2003). While the speed of development in large parts appears to be genetically determined, it was shown that A. bruennichi also exhibits great plasticity as a response to environmental cues (Cory & Schneider, 2018a), leading to faster growth or maturation in habitats with shorter warm seasons. The heritability of plasticity in A. bruennichi has so far not been studied in detail, though a degree of heritability is likely.
A. bruennichi has been described as an opportunistic predator of insects, observations in the field however, show orthopterans and hymenopterans to make up the greatest part of their diet (Bruggisser et al., 2012). As these insects rely on similar environmental factors regarding their development and reproduction, it is important for the spiders to adapt their development speed to the abundance of proportionately sized prey.
At different points of their life history, temperature, day length, and food availability play important roles for A. bruennichi’s life history. Our goal with this study was to experimentally determine how first-generation hybrid offspring of A. bruennichi are able to cope with these factors when compared to their parent populations. Additionally, we aimed to find out which adaptations of the hybrids might have enabled them to take advantage of the new ecological niche in Northern Europe.
We posited the following hypothesis regarding hybrid performance:
Hybrid offspring are better adapted to Northern European winter and summer conditions than their parent populations. We expected hybrid offspring to have higher survival rates during winter, exhibit faster growth during summer, and require less food during their growth period.
To test this hypothesis, we produced F1 offspring of two geographically distinct populations of A. bruennichi (both hybrid and pure-line offspring) and successively tested their performance under differing winter conditions (temperature) and summer conditions (temperature, day length, food availability).
Methods
Study Animals
The animals used in these experiments were offspring of spiders used in 2018 for experiments described in our previous study (Franke & Schneider, 2024). The parent generation consisted of 47 spiders from Tsukuba-Shi (Japan) and 55 spiders from Molleville (France). These were chosen as proxy for the Asian and Mediterranean ancestors of the naturally occurring hybrid population. Spiders mated in the lab and females were allowed to produce egg sacs until their natural death. This way, we produced both hybrid offspring (with one Asian and one Mediterranean parent), and pure line offspring (with both parents from Japan or France). Table 1 shows the amount of produced egg sacs from each of the four parental groups, as well as the number of matrilines.
Table 1 - Number of egg sacs produced by each of the four parental groups (FF: Mediterranean mother/Mediterranean father, FJ: Mediterranean mother/Asian father, JF: Asian mother/Mediterranean father, JJ: Asian mother/Asian father).
|
Parental Group |
Mother’s origin |
Father’s origin |
No. of egg sacs |
No of matrilines |
|
FF |
Mediterranean |
Mediterranean |
58 |
16 |
|
FJ |
Mediterranean |
Asian |
35 |
10 |
|
JF |
Asian |
Mediterranean |
40 |
13 |
|
JJ |
Asian |
Asian |
50 |
12 |
Experimental design
The egg sacs were laid between July and September 2018, transferred individually into acrylic boxes, and left under laboratory conditions (c. 24°C) until 2nd October 2018. We used sewing thread to suspend the egg sacs in the boxes, preventing contact to the container’s bottom. To test for the performance of the unhatched spiderlings, the egg sacs were then divided into three winter treatments (Germany, France, Japan), taking care to equally divide both the genetic groups and the matrilines among the three treatments. The egg sacs in the winter treatments France and Japan were transferred into a climate cabinet (RUMED 3501, Rubarth Apparate GmbH) with a climate program modelled after the average winter temperatures and day lengths in the original habitats of the parent population (Figure 1). The programs were set to reach their maximum temperature at noon, and then constantly decrease to their minimum at midnight, afterwards constantly increasing again. The temperature inside the cabinets was adjusted every other week. Since freezing temperatures are likely to be a developmental cue for A. bruennichi, as well as a selective pressure, we calculated the frost and ice days (days with lowest temperature below freezing and no temperature above freezing respectively) of each month (Table 2). We randomly included these frost and ice days for each month in our simulated French and Japanese climates. For frost days, we either substituted the average night temperature with 0°C (for France) or -1°C (for Japan) or left the night temperature unchanged if it would have been below freezing per our climate plan. To simulate ice days, we followed the same scheme for both day and night temperatures. Humidity was supplied in the form of a bowl of water placed in front of the cabinets’ ventilation fans, and sprayed water three times a week. The egg sacs in the German winter treatment were strung on thread and attached to wooden skewers. The skewers were placed outside in soil filled boxes. We left the egg sacs in the Germany treatment unattended for all of winter 2018/19. Temperatures used for the climate simulations, as well as frost and ice days, were derived from www.weatheronline.co.uk, using data averaged over 10 years (2004-2014). Day lengths were derived from www.timeanddate.com. For comparison with the simulated climates, we extracted the corresponding actual temperature values from www.meteostat.net for the local German winter conditions (Figure 1, Table 2).
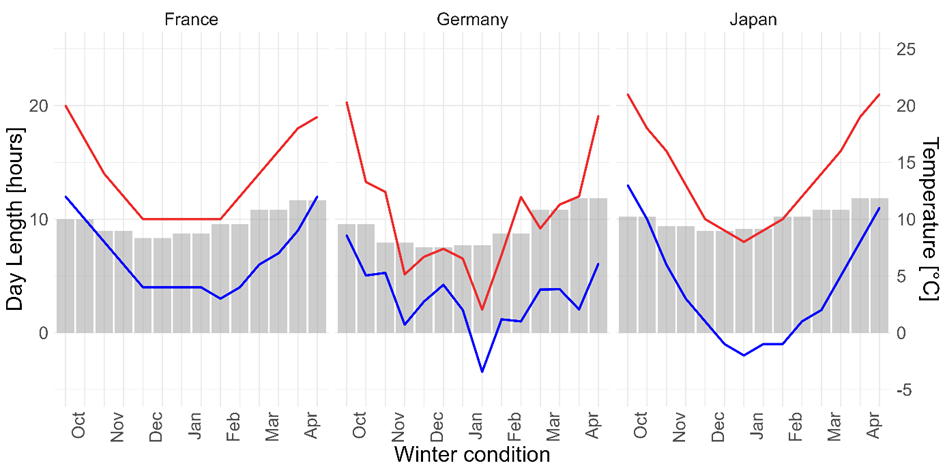
Figure 1 - Climate diagrams for the three overwintering treatments (France, Germany, Japan), red lines show highest day temperatures, blue lines lowest night temperatures, and bars the day length in hours. All values depict the values used in the climate simulations (for French and Japanese winter conditions), or the actual values for the German winter.
Table 2 - Frost days and ice days per month in the overwintering treatments.
|
France |
Oct |
Nov |
Dec |
Jan |
Feb |
Mar |
Apr |
|
Frost days |
0 |
1 |
5 |
5 |
5 |
0 |
0 |
|
Ice days |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Germany |
|||||||
|
Frost days |
1 |
5 |
5 |
10 |
13 |
2 |
6 |
|
Ice days |
0 |
0 |
0 |
4 |
0 |
0 |
0 |
|
Japan |
|||||||
|
Frost days |
0 |
1 |
23 |
31 |
22 |
2 |
0 |
|
Ice days |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
On the 3rd of May 2019, we transferred the egg sacs in their plastic containers into two walk-in climate chambers (Weiss Technik, WK 21’/5-40). The climate chambers were programmed to simulate the mean temperature and day length during spring and summer in the origin location in France, as well as local conditions in Hamburg for the German climate treatment. Temperatures were adjusted to reach the highest day temperature at noon and the lowest night temperature at midnight, rising and falling over the day. We adjusted the temperature and day length at the beginning of each subsequent week (Figure 2). The egg sacs were misted with water five times a week. Again, we took care to evenly distribute the parental groups, as well as the winter treatments among the two climate chambers.
After about a week we carefully opened the egg sacs with microscopy scissors to control for hatching. We noted which egg sacs contained hatched individuals, and which did not. We waited for the spiderlings to leave the egg sacs and allowed them to build their communal web inside the plastic containers. Once in these communal webs, we transferred a total of 15 spiders from each egg sac into individual upturned plastic cups (40 mL). We subsequently adjusted the cup size in accordance with the growth of the spiders (200 mL; 500 mL). In the case of a lower number of total egg sacs for a given treatment, we isolated more than 15 individuals to match the numbers. This was especially the case for those with a French mother and Japanese father (FJ), as there was a lower hatching rate for this combination and particularly few spiderlings hatched from egg sacs that overwintered under French and Japanese conditions. A total of 2869 individual spiders were isolated and used for analyses.
Spiders were watered five to six times a week by spraying their webs and directly soaking a piece of cotton inside the housing cup above the spider. They were fed twice weekly with Drosophila melanogaster or D. hydei. We introduced two feeding treatments: spiderlings in the High Food treatment were fed ad libitum and spiderlings in the Low Food treatment were fed the same number of flies but their cups were lifted the following day so that the remaining flies alive were removed from the cups, leading to a reduced food availability.
We checked each spider six days of the week for moulting or death and recorded the dates for these events. Mortality rates were very high during our experiments, resulting in our inability to analyse any influences of the treatments on adult weight, or maturity rates. We could however analyse the survival rates until 10th October (last day of data collection), as well as the developmental state at this date. Additionally, we calculated the mean number of moulting events and days between moulting events. The first moult of A. bruennichi typically occurs inside the egg sac, so we were able to note all subsequent moults of the isolated spiderlings. As mortality was high, we analysed growth up to the 4th moult, which was reached by a sufficiently large number of individuals.
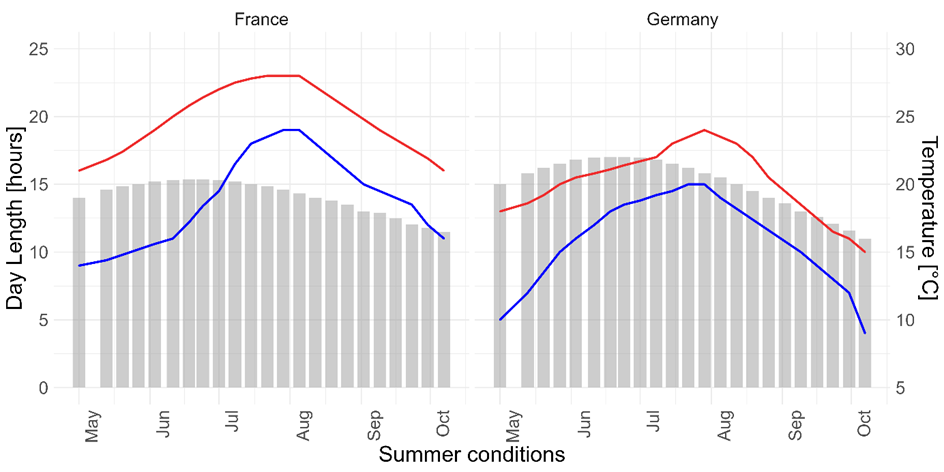
Figure 2 - Climate diagrams for the two summer treatments (France, Germany), red lines show highest day temperatures, blue lines lowest night temperatures, and bars the day length in hours. All values depict the actual values of the climate simulations.
Statistical Analysis
We performed statistical analysis using R Studio (R version 4.5.1).
We analysed hatching success using generalized linear mixed models (GLMM) with a binomial error distribution. We used the R package glmmTB, as this framework provides more robust estimates for binomial GLMMs. Fixed effects were winter condition (France/Germany/Japan) and genetic combination (FF, FJ, JF, JJ), and the interaction of both factors, with random intercepts for matriline and egg sac number. Post hoc comparisons were conducted by using the package emmeans, with corrections for p-values according to the Tukey HSD method.
To compare performance between the pure line and hybrid offspring, we focussed on mortality and growth of individuals during the second experiment. We analysed both mortality (time from separation to death) and growth (time from separation to fourth moult) using Cox proportional hazards mixed-effects models (R package coxme), with food treatment (High/Low), climate treatment (France/Germany) and genetic combination as fixed effects and matriline and egg sac number as random intercepts to account for non-independence. We tested the proportional hazards (PH) assumption for the models using Schoenfeld residual tests, revealing violations for several variables. We still decided to proceed with the standard Cox model due to our large sample size (> 2000), which can result in the detection of minor proportionality departures that may not be practically meaningful. Additionally, a recent study found only a negligible impact of PH violations on model calibration, compared to alternative models (Austin & Giardiello, 2025). We adjusted p-values derived from our final models using the Benjamini-Hochmann correction method for multiple testing. We additionally fitted equivalent fixed-effect models using the coxph function to obtain estimated marginal means and contrasts for fixed effects with more than two levels via the emmeans package, with p-values adjusted using the Tukey HSD method.
Model simplification in all analyses was performed using a backwards elimination approach, based on likelihood ratio tests by sequentially removing non-significant interaction terms from the full model. The results of the full Cox proportional hazards models before simplification can be found in the appendix. Significant results of the fully reduced models are reported as hazard ratios (HR) with 95% confidence intervals and p-values.
Results
Hatching success
Hatching success was defined as the proportion of egg sacs to contain viable spiderlings, with the response being either success or no success. We were unable to determine the exact number of eggs contained in the individual egg sacs, as often unhatched eggs were fused together inside the concrete-like centre of the egg sac, which prevented us from exactly counting the amount. Egg sacs had no hatching success when all spiderlings were dead or deformed, or when only a very small number of spiderlings (< 20) out of the hundreds of eggs laid were alive and viable. We analysed hatching success of the first to seventh egg sacs of all matrilines. While we found differences in the hatching success between the winter climates (France: 12.3% ± 8.7, Germany: 26.3% ± 14.6, Japan: 29.9 ± 15.7), these were not significant (LRT: Χ² = 5.47, d.f. = 2, p = 0.07); neither were effects of the interaction of winter condition and genetic combination (LRT: Χ² = 4.05, d.f. = 6, p = 0.67). However, genetic combination had a strong effect (LRT: Χ² = 9.99, d.f. = 3, p = 0.019; Figure 3), with FJ clutches showing markedly lower hatching success (4.3% ± 4.3) than JF (42.1% ± 21.7, p = 0.048) and JJ (42.6% ± 20.5, p = 0.037). Egg sacs from the FF background meanwhile were not different from the other groups (19.9% ± 13.2).
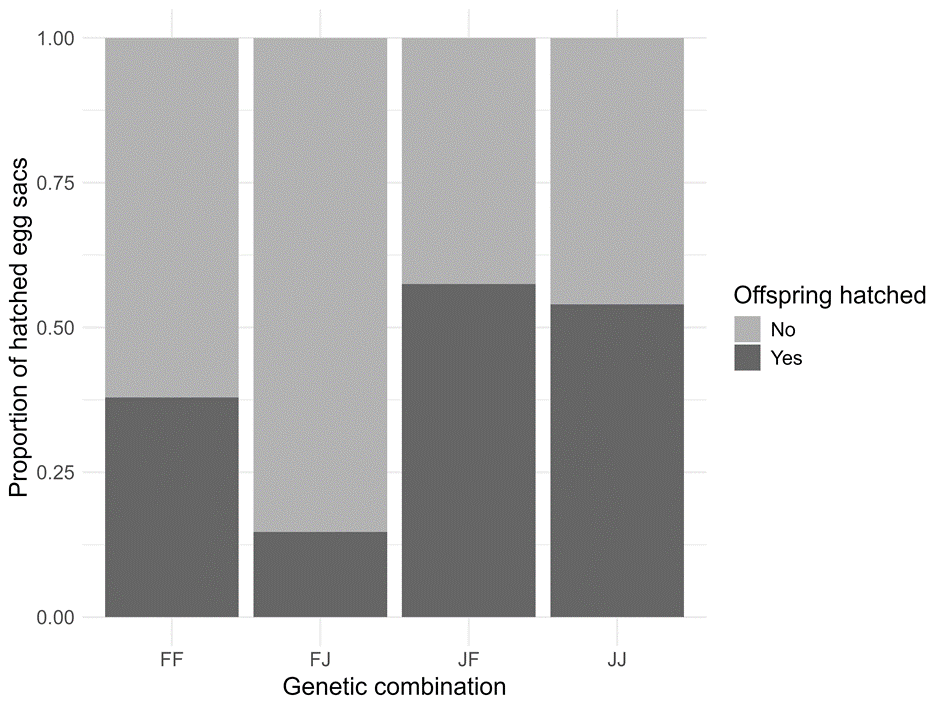
Figure 3 - Proportion of egg sacs successfully hatched of the four different genetic combinations (FF: Mediterranean mother/Mediterranean father, FJ: Mediterranean mother/Asian father, JF: Asian mother/Mediterranean father, JJ: Asian mother/Asian father) pooled over three winter climate treatments.
Hybrid performance
Mortality
Mortality was measured in total days alive after separation into individual containers and showed variation between the genetic combinations (FF: 95.6 ± 24.1, FJ: 80.9 ± 35.1, JF: 96.5 ± 27.5, JJ: 95.5 ± 26.8), as well as between climate (France: 96.7 ± 26.6, Germany: 92.1 ± 28.4) and food treatments (High: 95.1 ± 27.4, Low: 93.8 ± 27.8). In total, our analysis included 2638 spiders, of which 2429 had died by experiment end.
The effect on mortality from the interaction of the food treatment and genetic background proved non-significant (LRT: Χ² = 7.28, d.f. = 3, p = 0.063) but revealed a trend. Post-hoc analyses, however, revealed that survival differences among genetic groups were dependent on the food availability. To avoid masking these potentially biologically relevant effects, the interaction term was retained in the final model, and results will be given here.
We found that the food treatment had a significant effect only on the survival of the JF combination, with higher survival under high than low food availability (HR = 0.83 (95% CI: 0.72-0.95), p = 0.007), an effect not observed in all other genetic groups. Comparing the performance of the genetic groups within food treatments, we saw that the FJ combination showed significantly worse survival than the JF combination under high food conditions (HR = 1.36 (95% CI: 1.11-1.68), p = 0.018), and both the FF (HR = 1.37 (95% CI: 1.11-1.69), p = 0.018) and JJ (HR = 1.40 (95% CI: 1.14-1.73), p = 0.009) under low food conditions (Figure 4).
There also was a strong influence on mortality of the climate condition (HR = 1.14 (95% CI: 1.05-1.24), p = 0.003), with the German climate leading to significantly earlier death than the French climate.
We did not find effects on survival from the interaction of genetic combination and climate (LRT: Χ² = 4.38, d.f. = 3, p = 0.22), the interaction of the food and climate treatment (LRT: Χ² = 1.87, d.f. = 1, p = 0.17), nor the three-way interaction (LRT: Χ² = 2.39, d.f. = 1, p = 0.49).
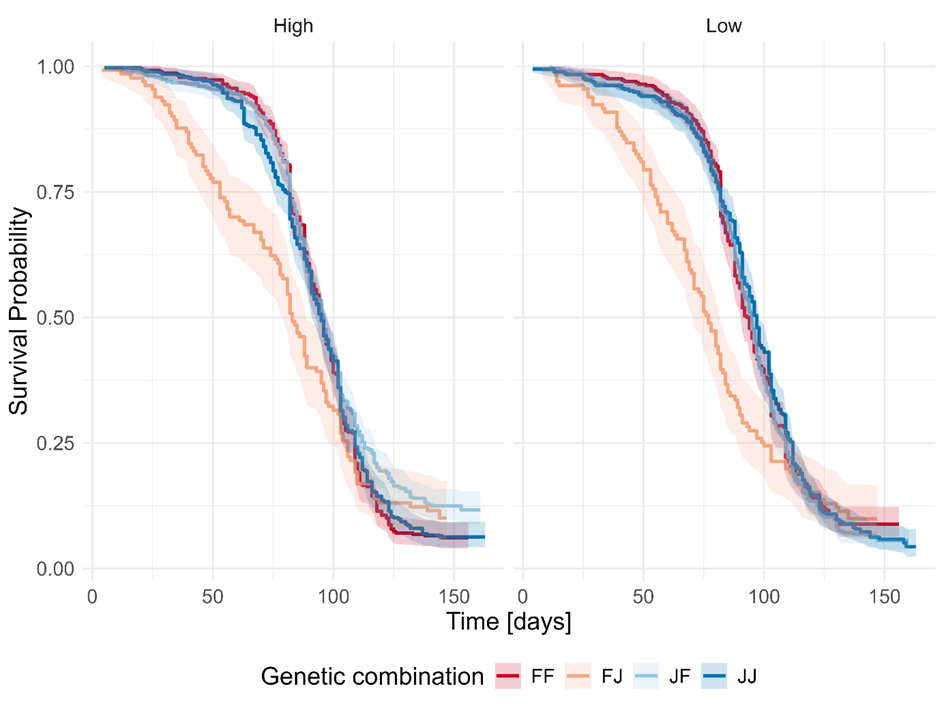
Figure 4 - Kaplan Meier curves describing survival probability for the four genetic combinations (FF: Mediterranean mother/Mediterranean father, FJ: Mediterranean mother/Asian father, JF: Asian mother/Mediterranean father, JJ: Asian mother/Asian father) in the two food treatments (High and Low), pooled over the climate treatments.
Growth
Growth was defined as the days needed to reach the fourth moult, with 1978 of 2080 analysed spiders achieving this event. We observed notable differences in the growth time between the genetic combinations (FF: 79.6 ± 19.0, FJ: 90.9 ± 37.0, JF: 82.5 ± 22.5, JJ: 85.2 ± 23.6), the climate treatment (France: 86.3 ± 21.7, Germany: 79.1 ± 23.3), and the food treatment (High: 80.9 ± 22.9, Low: 84.8 ± 22.4).
Contrary to the analysis of mortality, we found that growth was significantly affected by an interaction between the genetic combination and the climate treatment (LRT: Χ² = 9.86, d.f. = 3, p = 0.019). The German climate led to consistently faster growth than the French climate (Figure 5), with the effect being significant for the combinations FJ (HR = 0.48 (95% CI: 0.27-0.84, p = 0.011), JF (HR = 0.68 (95% CI: 0.59-0.79, p < 0.0001), and JJ (HR = 0.78 (95% CI: 0.66-0.91, p = 0.003), as well as showing a trend in the FF combination (HR = 0.86 (95% CI: 0.73-1.01), p = 0.06). In the French climate we saw spiders with an FF background exhibiting faster growth than all other groups (FJ: HR = 3.13 (95% CI: 1.89-5.18), p = 0.0001, JF: HR = 1.27 (95% CI: 1.09-1.47), p = 0.009, JJ: HR = 1.39 (95% CI: 1.18-1.63), p = 0.0004). On the other hand, spiders with the hybrid background FJ showed slower growth than all other groups (JF: HR = 0.41 (95% CI: 0.25-0.67), p = 0.002, JJ: HR = 0.44 (95% CI: 0.28-0.73), p = 0.009). In the German climate FJ grew slower than the pure Mediterranean FF (HR = 0.57 (95% CI: 0.42-0.78), p = 0.002), and the other hybrids JF (HR = 0.58 (95% CI: 0.43-0.78, p = 0.002). Here, the pure Asian JJ exhibited a slower growth rate than spiders with both the FF (HR = 0.80 (95% CI: 0.68-0.94), p = 0.033) and JF backgrounds (HR = 0.80 (95% CI: 0.68-0.94), p = 0.032).
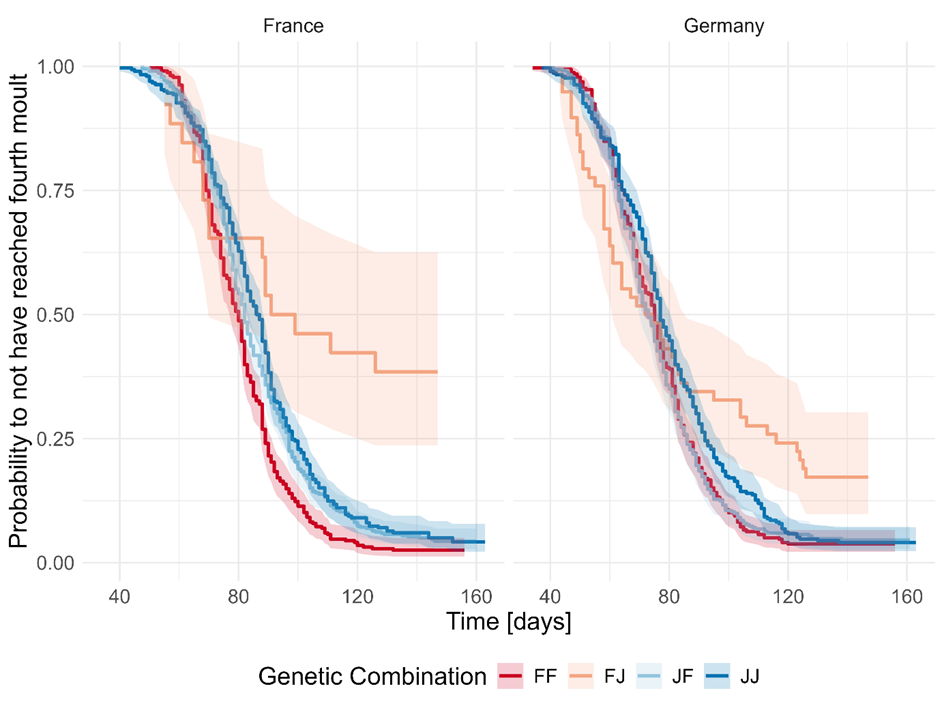
Figure 5 - Kaplan Meier curves describing probability of not having reached the fourth moult for the four genetic combinations (FF: Mediterranean mother/Mediterranean father, FJ: Mediterranean mother/Asian father, JF: Asian mother/Mediterranean father, JJ: Asian mother/Asian father) in the two climate treatments (France and Germany), pooled over the food treatments.
The food treatment also showed a highly significant effect on growth (HR = 0.82 (95% CI: 0.75-0.89), p < 0.001), with spiders reaching their fourth moult later under low food availability.
We could not find any effects of the interactions of food and climate (LRT: Χ² = 0.23, d.f. = 1, p = 0.63), and food and genetic combination (LRT: Χ² = 3.56, d.f. = 3, p = 0.31), nor the three-way interaction of the three fixed effects (LRT: Χ² = 1.12, d.f. = 3, p = 0.77).
Discussion
We conducted a large breeding study crossing Argiope bruennichi spiders from Mediterranean and Asian populations to test the hypothesis that hybrids would perform better under challenging conditions than pure line offspring due to hybrid vigour. We could show that offspring from Mediterranean mothers and Asian fathers (FJ), compared to the other genetic combinations, had significantly lower hatching success after overwintering, as well as showing lower survival and slower growth during spring and summer. Hybrid spiders with an Asian father and a Mediterranean mother (JF) on the other hand performed similarly to their parent populations. JF hybrids even exhibited faster growth in the challenging German climate than their pure Asian parent line. Their survival however was negatively affected by low food availability, which was not observed in the other lines.
Our predictions were not confirmed by our experimental results. We predicted that hybrids would outcompete the pure line in their hatching rate, and their survival and growth rates under German climate and lower food availability. Contrary to our expectations, one hybrid line (FJ) performed worse than all other groups, while the other (JF) performed comparably to the pure lines. Overall, FJ hybrids were affected during all stages, while JF hybrids only showed differences during the growth period.
The results for the FJ genetic combination are well in line with genetic incompatibility and outbreeding depression. The combination of alleles that these hybrids received via recombination were apparently detrimental to their hatching success, with their hatching rate less than half that of the other groups. It is possible that responsible alleles could be relevant for cold tolerance or resource allocation. These traits are known to play an important role for A. bruennichi and show considerable population differences (Sheffer et al., 2024). Detrimental combinations would have led to either freezing or higher than optimal metabolic rates that would have left spiders too weak to hatch after winter. Such effects could also have contributed to the decreased survival and growth speed of this genetic group during summer, resulting in the observed differences. Combined, low hatching success and stunted performance represent an effective post-zygotic barrier to hybridisation of these specific pairings, in a spider that has no pre-zygotic isolation mechanisms (Franke & Schneider, 2024).
A general incompatibility of Asian and Mediterranean populations can however be ruled out by the performance of the other hybrid group (JF). Hatching success for these spiders showed no differences from the parent groups, and while they were the only combination negatively affected by food shortage in their survival, they managed to outperform pure Asian spiders in growth speed. This lets us conclude that the genetic combination had similar adaptations at their disposal regarding traits connected to overwintering and growth. The asymmetric performance of the two hybrid groups is indicative of incompatibilities relating to sex-heritable loci, which would be found on sex chromosomes or mtDNA. Since spiders have an XX/X0 sex determination system, mitonuclear incompatibility appears to be the most likely to affect this species. This phenomenon is a well-established driver of asymmetric hybrid breakdown, with recent studies in various taxa from copepods to birds identifying mitonuclear mismatches as a primary cause of reduced hybrid fitness (Healy & Burton, 2023; Shogren et al., 2024). In this case, mtDNA transferred from Mediterranean mothers into the hybrids would not be able to properly interact with the nuclear-encoded genome transferred by their Asian fathers, which would be adapted to function with Asian mtDNA. Considering the importance of mtDNA-encoded loci for energy production inside the cell, any incompatibilities would result in reduced viability, as we have observed in our study (Burton, 2022). As our study only looked at life history of the hybrids, the underlying mechanism of incompatibility would have to be explored by further genetic studies with an additional focus on introgression of mtDNA.
The observed asymmetry between the two hybrid groups confirms the high importance of the right genetic combinations for a hybridisation event. This “Darwin’s Corollary”, where reciprocal hybrids exhibit different fitness, is a widely observed phenomenon in animal systems (Schneemann et al., 2022). Such directional, asymmetric outcomes are often the result of complex interactions that can lead to outbreeding depression in one cross, but not the other. In our case we did not observe any hybrid vigour. It is however possible that other life history traits such as maturation speed or fertility and fecundity could show signs of hybrid vigour. Our results suggest that offspring of Mediterranean mothers and Asian fathers would contribute only little to the gene pool of a hybridising population under natural conditions, as their significant mortality during both winter and summer would have rendered a large proportion inviable. In contrast, hybrids from Asian mothers and Mediterranean fathers would mostly be able to compete with the two parental populations, and contribute significantly to the gene pool, thus facilitating the process towards hybrid stabilization.
These results contrast those from our previous study, which was conducted on the parents of both our hybrid and pure line spiders (Franke and Schneider 2024). In that behavioural study we focussed on the complex mating system of these spiders, and found significant indications of directed hybridisation, with the FJ combination being more prone to contribute to forming a hybrid population. While the combined results from both studies don’t allow us to pinpoint the specific combination that contributed the most to the Northern European hybrids, they do not contradict each other. We only tested F1 hybrids, and while F1 studies provide important initial insights, understanding the long-term evolutionary potential often requires examining F2 and later generations. Commonly, hybrid vigour is expressed only in the F2 generation, often even after an expression of outbreeding depression, due to ongoing recombination in later generations and thus potentially different fitness outcomes (Edmands, 2007; Schneemann & Welch, 2025). Repeating this study with F2 hybrids, as well as repeating the behavioural assays with F1 hybrids, could reveal further information about this interesting system.
It is also important to note that the French and Japanese spiders we used very likely do not represent the actual historic ancestry populations of the Northern European hybrids. More probably, Mediterranean individuals closer to the northern hybrid zone contributed to the new populations, such as Northern France or Southwest Germany. The likely origin of the Asian introgression into the hybrid population has been suggested to lie in Central Asia or European Russia (Krehenwinkel et al., 2015), although Japanese spiders have also been used for the genetic analyses in lieu of Central Asian individuals. Thus, the specific genetic makeup of our experimental parental populations may differ from the actual founding populations involved in the historical range expansion. Another layer of complexity stems from the inherently random nature of genetic recombination. Allele combinations inherited by our experimental F1 hybrids only represent one possible outcome, the observed outbreeding depression in the FJ combination and general lack of hybrid vigour. The historical hybridisation event likely involved various founding individuals and sets of alleles, which could have, by chance, resulted in more advantageous allele combinations than those observed in our study.
Our results regarding the general influences of the climate and food treatments suggest that we were successful in simulating adverse conditions which affected individual performance. As expected, food availability strongly influenced growth speed, as food shortage would have resulted in resources being locked in metabolic maintenance being unavailable for growth allocation. Our climate treatment too affected survival and growth across genetic combinations, with the growth patterns especially warranting closer discussion. Spiders of all backgrounds reached their fourth moult faster when exposed to the German climate conditions, a clear sign of phenotypic plasticity, a key mechanism by which organisms can cope with rapid environmental change (Stollewerk et al., 2025). This pattern is consistent with counter-gradient variation, where populations from higher latitudes with shorter seasons evolve faster intrinsic growth rates to ensure timely maturation (Gotthard et al., 2025). Because our climate treatment varied both temperature and day length profiles, we cannot definitively isolate the specific trigger for this accelerated growth. However, since lower temperatures typically slow developmental rates in ectotherms, the observed acceleration strongly suggests that photoperiod, or the interaction between photoperiod and temperature, plays a crucial role as an environmental cue. Such rapid adaptation of photoperiodic response is a well-documented mechanism in arthropods, aiding in range expansion (Urbanski et al., 2012; Smith et al., 2021; Neu & Fischer, 2022). Further experiments manipulating temperature and day length independently would be necessary to fully disentangle their respective contributions, as has been explored in systems like damselflies from different latitudinal populations (Sniegula et al., 2013). As was shown in other studies(Cory & Schneider, 2018b, 2018a), A. bruennichi from Germany synchronise their maturation more strongly than spiders from Southern France, thereby ensuring sufficient available mating partners. The observed higher mortality under German climate can likely also be attributed to this increased plastic growth speed, as faster growth can be stressful for individuals and requires shorter intervals between moults. This suggests a potential cost or constraint to this plastic adaptation. A pattern where plastic developmental responses occur but are nonetheless restricted by high mortality from other climate stressors was recently documented in range-expanding butterflies (Ittonen et al., 2025) and shows similarities to our system. The overarching similar patterns in growth suggest a high degree of plasticity for A. bruennichi across geographical populations, a trait very likely facilitating the colonisation of novel environments such as Central and Northern Europe.
The hybrid offspring we studied in our experiment exhibited a strongly asymmetric performance, depending on the geographical origin of their parents. We found strong indications for mitonuclear conflict in our system, resulting in F1 hybrids with either extreme outbreeding depression, or comparable performance. In addition, we could show that A. bruennichi possesses developmental plasticity regarding growth, with temperature, day length, or a combination being likely triggers for development.
Appendices
Appendix 1 - Initial distribution of spiders in the different treatment groups of the hybrid performance experiment. A total of 2868 spiders were analysed.
|
Genetic Combination |
Winter Conditions |
Spring and Summer Conditions |
Food Treatment |
No. of Spiders |
No. of Matrilines |
|
FF |
Germany |
Germany |
High |
90 |
6 |
|
Low |
90 |
6 |
|||
|
France |
High |
90 |
6 |
||
|
Low |
90 |
6 |
|||
|
France |
Germany |
High |
47 |
3 |
|
|
Low |
47 |
4 |
|||
|
France |
High |
45 |
3 |
||
|
Low |
45 |
3 |
|||
|
Japan |
Germany |
High |
60 |
4 |
|
|
Low |
61 |
4 |
|||
|
France |
High |
61 |
4 |
||
|
Low |
60 |
4 |
|||
|
FJ |
Germany |
Germany |
High |
75 |
3 |
|
Low |
75 |
3 |
|||
|
France |
High |
60 |
3 |
||
|
Low |
60 |
3 |
|||
|
France |
Germany |
High |
/ |
/ |
|
|
Low |
/ |
/ |
|||
|
France |
High |
/ |
/ |
||
|
Low |
/ |
/ |
|||
|
Japan |
Germany |
High |
15 |
3 |
|
|
Low |
15 |
3 |
|||
|
France |
High |
/ |
/ |
||
|
Low |
/ |
/ |
|||
|
JF |
Germany |
Germany |
High |
61 |
4 |
|
Low |
61 |
4 |
|||
|
France |
High |
60 |
4 |
||
|
Low |
60 |
4 |
|||
|
France |
Germany |
High |
60 |
4 |
|
|
Low |
60 |
4 |
|||
|
France |
High |
92 |
5 |
||
|
Low |
94 |
5 |
|||
|
Japan |
Germany |
High |
105 |
7 |
|
|
Low |
106 |
6 |
|||
|
France |
High |
107 |
6 |
||
|
Low |
105 |
6 |
|||
|
JJ |
Germany |
Germany |
High |
60 |
4 |
|
Low |
60 |
4 |
|||
|
France |
High |
60 |
4 |
||
|
Low |
61 |
4 |
|||
|
France |
Germany |
High |
76 |
5 |
|
|
Low |
76 |
5 |
|||
|
France |
High |
71 |
5 |
||
|
Low |
73 |
5 |
|||
|
Japan |
Germany |
High |
62 |
4 |
|
|
Low |
61 |
4 |
|||
|
France |
High |
76 |
3 |
||
|
Low |
75 |
3 |
Appendix 2 - Results of full Cox Proportional Hazards models analysing mortality and growth
|
Mortality |
|||
|
Fixed effects |
Hazard ratio (95% CI) |
z-value |
p-value |
|
Food |
0.90 (0.73-1.11) |
-0.97 |
0.33 |
|
Climate |
0.93 (0.75-1.15) |
-0.70 |
0.48 |
|
Genetic combination FJ |
0.96 (0.64-1.44) |
-0.18 |
0.86 |
|
Genetic combination JF |
0.83 (0.64-1.07) |
-1.45 |
0.15 |
|
Genetic combination JJ |
0.92 (0.70-1.21) |
-0.57 |
0.57 |
|
Food : Climate |
1.15 (0.85-1.56) |
0.93 |
0.35 |
|
Food : Genetic combination FJ |
1.49 (0.93-2.37) |
1.67 |
0.09 |
|
Food : Genetic combination JF |
1.21 (0.91-1.61) |
1.34 |
0.18 |
|
Food : Genetic combination JJ |
1.02 (0.76-1.38) |
0.14 |
0.89 |
|
Climate : Genetic combination FJ |
1.62 (1.02-2.56) |
2.04 |
0.042 |
|
Climate : Genetic combination JF |
1.14 (0.86-1.53) |
0.91 |
0.36 |
|
Climate : Genetic combination JJ |
1.26 (0.93-1.69) |
1.50 |
0.13 |
|
Food : Climate : Genetic combination FJ |
0.68 (0.37-1.26) |
-1.23 |
0.22 |
|
Food : Climate : Genetic combination JF |
1.08 (0.71-1.62) |
0.35 |
0.73 |
|
Food : Climate : Genetic combination JJ |
0.92 (0.60-1.40) |
-0.39 |
0.69 |
|
Growth |
|||
|
Food |
0.69 (0.55-0.87) |
-3.20 |
0.001 |
|
Climate |
1.09 (0.88-1.37) |
0.82 |
0.41 |
|
Genetic combination FJ |
0.21 (0.09-0.49) |
-3.65 |
0.0003 |
|
Genetic combination JF |
0.83 (0.56-1.22) |
-0.97 |
0.33 |
|
Genetic combination JJ |
0.72 (0.47-1.08) |
-1.58 |
0.11 |
|
Food : Climate |
1.11 (0.81-1.53) |
0.65 |
0.52 |
|
Food : Genetic combination FJ |
1.88 (0.66-5.31) |
1.19 |
0.23 |
|
Food : Genetic combination JF |
1.17 (0.86-1.57) |
1.00 |
0.32 |
|
Food : Genetic combination JJ |
1.31 (0.95-1.81) |
1.62 |
0.10 |
|
Climate : Genetic combination FJ |
2.69 (1.14-6.38) |
2.25 |
0.024 |
|
Climate : Genetic combination JF |
1.31 (0.97-1.76) |
1.75 |
0.08 |
|
Climate : Genetic combination JJ |
1.28 (0.93-1.77) |
1.50 |
0.13 |
|
Food : Climate : Genetic combination FJ |
0.68 (0.20-2.29) |
-0.63 |
0.53 |
|
Food : Climate : Genetic combination JF |
0.99 (0.64-1.53) |
-0.05 |
0.96 |
|
Food : Climate : Genetic combination JJ |
0.83 (0.52-1-31) |
-0.80 |
0.43 |
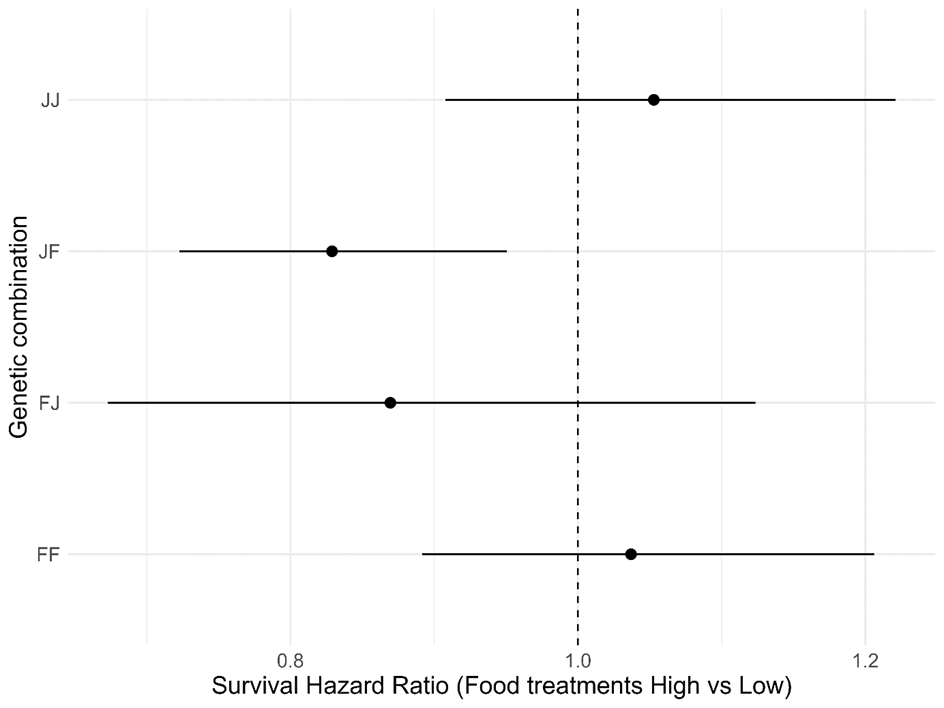
Appendix 3 - Hazard ratios for the effect of the food treatments (High, Low) on the survival of the four genetic combinations (FF, FJ, JF, JJ). A significant effect is visible for JF, which survived longer under high than under low food conditions.
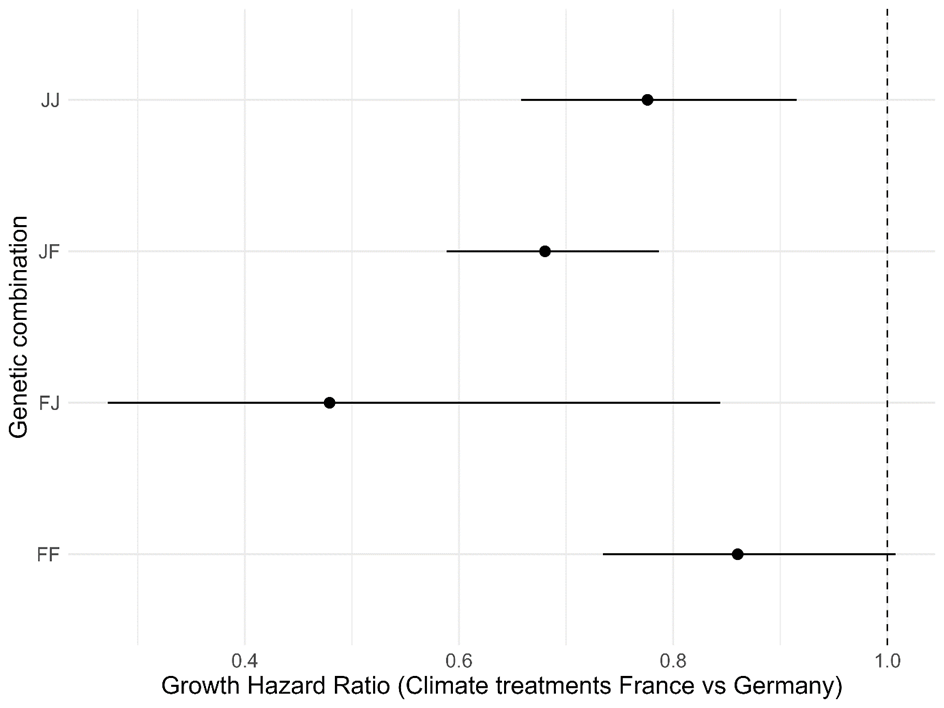
Appendix 4 - Hazard ratios for the effect of the climate treatments (France, Germany) on the growth of the four genetic combinations (FF, FJ, JF, JJ). Significant effects are visible for the backgrounds FJ, JF, and JJ, which grew faster in German than in French climate.
Acknowledgements
We are very grateful for the help of the people who collected the parent spiders of offspring used in our experiments: Anna-Lean Cory and Claudia Drees in France, and Yuki Baba in Japan; as well as the host of helpers without whom care of the spiders would not have been possible: Our technicians Angelika Taebel-Hellwig, Tomma Dirks, Jasmin Kriske, and student helpers Michelle, Wilma, David, and Jessika. We also want to thank Aimee Deconinck, Matthias Foellmer and Raphaël Royauté, as well as an anonymous reviewer for their helpful comments and suggestions. Preprint version 2 of this article has been peer-reviewed and recommended by PCI Evol Biol (https://doi.org/10.24072/pci.evolbiol.100870; Jourdan-Pineau, 2025).
Data, scripts, code, and supplementary information availability
Data and scripts are available online (https://doi.org/10.17605/OSF.IO/3JFM5; Franke et al. 2025).
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Funding
All work was conducted in the framework of the project ‘Hybrids – Chances and Challenges of New Genetic Combinations’, University of Hamburg.