
 CC-BY 4.0
CC-BY 4.0
Introduction
Seagrass habitats play a key role in coastal ecosystems, representing local hotspots of species richness (Kindeberg et al., 2022; Pierrejean et al., 2023) and stimulating secondary production in, and around the area they colonise (Rodil et al., 2020). Seagrasses have been highlighted for their potential role in reducing surface seawater CO2 concentrations - thereby locally mitigating ocean acidification (e.g. Hendriks et al.; 2014; Ricart et al., 2021) - as well as in carbon storage and sequestration (Röhr et al., 2018; Christensen 2023), due to high rates of biological production and biomass accumulation. Healthy seagrass habitats tend to be net autotrophs due to an efficient photosynthesis/respiration ratio and are therefore efficient carbon sinks (Duarte and Cebrián, 1996; Gattuso et al., 1998). Conversely, a severe loss in seagrass shoot density (of 50–60%) has been shown to shift the entire habitat towards heterotrophy, due to decreased photosynthesis and respiration caused by lowered biomass, as well as increased microbial activity in the dead and decomposing biomass (Berger et al., 2020).
Over the last few decades, eutrophication, mechanical disturbance and climate change have been the main causes of seagrass decline worldwide (e.g. Orth et al., 2006). Some promising positive reversal trends have been observed in Europe, where management has created opportunities for recovery (Dunic et al., 2021). However, this vulnerability raises questions about the role these habitats play in both the overall functioning of coastal ecosystems, particularly with regard to carbon cycling. Empirical assessments of community metabolism are therefore essential to quantify the potential net effect of seagrass meadows of varying environmental states on carbon budgets. Accurate assessment of benthic metabolism in seagrass communities is however challenging, due to both the logistical complexity of the processes to be measured and their natural variability in time and space (Macreadie et al., 2014). Metabolism is highly mutable and dependant on a complex set of factors such as species, local biotic interactions, local hydrodynamics and associated physical characteristics, all of which vary seasonally and geographically (Ward et al., 2022). Temporal changes in seagrass morphometric traits and in associated species assemblages are also local drivers of seagrass metabolism over time (Delgard et al., 2016; Rodil et al., 2022).
Comparative data using the same methodology to describe variations in seagrass metabolism at multiple scales is still lacking. Temporal studies are often site-specific (Yarbro and Carlson, 2008; Ouisse et al., 2010; Champenois and Borges, 2012), whereas spatial variability is often reduced to season-specific approaches (Asmala et al., 2019; Bravo and Grant, 2020). Moreover, most of the studies are performed in spatially stable and homogeneous meadows, while meadows are known to be highly dynamic, with peripheral areas subject to more variations (a.k.a. the “edge effect”, Hovel and Lipcius, 2001; Turrietta and Reidenbach, 2024), and adjacent bare areas influenced by seagrass proximity (Muller et al., 2023). Few studies have evidenced an edge effect on benthic fluxes in Cymodocea nodosa (Barrón et al., 2006) or Zostera noltii (Delgard et al., 2016) habitats. The metabolic capacities of seagrass meadows are also influenced by local features and environmental spatial gradients within the seagrass beds themselves (Ricart et al., 2020).
This study aimed to investigate the variability in seagrass community benthic metabolism during immersion within the distribution range of two seagrass species in France, Zostera marina Linnaeus, 1753, and Zostera noltii Hornemann, 1832, while accounting for local meadow-scale fluctuations in these habitats. Three areas were investigated: the core and edge of the meadow as well as adjacent bare sediment. Seasonal variations were considered by conducting sampling at five different time periods over two years. We measured both oxygen (O2) and dissolved inorganic carbon (DIC) fluxes at the community level i.e by considering seagrasses and associated species.
Among benthic processes, primary production is often derived from O2 measurements, which are more accurate and easier to obtain continuously than carbon measurements. To infer about carbon cycling, O2 values are then converted to carbon using the photosynthetic quotient, that is the molar ratio of the flux of O2 released to the CO2 assimilated during photosynthesis. A theoretical ratio of 1:1 is commonly used, without appreciating the broad range of possible values (Trentman et al., 2023). This study also aimed to improve estimates of carbon conversion based on O₂ production by conducting simultaneous O₂ and DIC measurements.
To the best of our knowledge, this study is the first to deploy standardised in situ measurements, while factoring in seasonal effect at different sites and habitats, varying both in terms of macrophyte cover (seagrass vs bare) and meadow heterogeneity (stable vs dynamic area seagrass bed areas). Our results will help to address the context-dependency of spatio-temporal fluctuations in seagrass metabolism, which is essential for effectively incorporating seagrass habitats into broader-scale modelling efforts. Ultimately, these efforts are critical to scale up important ecosystem functions such as carbon cycle, and propose future scenarios, particularly in the context of global change.
Materials and methods
Study sites and sampling strategy
The functional role of seagrass during immersion was investigated by measuring DIC and O2 fluxes at the sediment-water interface in Z. marina and Z. noltii habitats and adjacent bare areas at four sites across the seagrass habitat distribution range along the French coasts (Dinard, Gulf of Morbihan, Bay of Arcachon and Thau lagoon, Figure 1). One important difference between the sites is the decreasing gradient in tidal amplitude from the northernmost Dinard site to the southernmost site, with the tide being negligible at Thau. Thus, both Z. marina and Z. noltii are always submerged at Thau whereas Z. noltii is intertidal at other sites. The maximum temperatures also illustrate the north-south distribution gradient of the sites (Table 1).
Nine benthic chambers were deployed seasonally by SCUBA-diving in each habitat (Z. marina and Z. noltii, Figure 1) at the four sites between winter 2020 and spring 2021 (winter, summer, autumn 2020 and winter and spring 2021). At each site, three spatio-temporal modalities were defined for each Zostera habitat, as gathered from historical maps and/or area photographs where available, or based on local expert knowledge: (1) a “core” area of the meadow where historic spatial data showed a stable meadow occurrence through time (at least 10 years) (S modality), (2) a dynamic area of the meadow where fluctuating seagrass presence and absence were observed over short alternating periods (D modality, not identified in Arcachon), and (3) an “absent” constantly bare area adjacent to meadows where no seagrass has ever been observed (A modality) (Lacoste et al., 2024). Fluxes were measured in triplicate in each of these three discrete areas.
Table 1 - Location, annual water temperature variations, tidal regime and bed size at the four study sites. Data retrieved from Lacoste et al., (2025).
Site | Coordinates (meadow core, longitude; latitude) | Bed size (ha) | Tidal regime | Water temperature range* (°C) | ||
|---|---|---|---|---|---|---|
Z. marina | Z. noltii | Z. marina | Z. noltii | |||
Dinard | -2.04367; 48.63394 | -2.19814; 48.60327 | 217 | 27 | Mega | 10-17.4 |
Morbihan | -2.75789; 47.56318 | -2.7552; 47.54044 | 812 | 544 | Meso | 9.7-18.6 |
Arcachon | -1.18967; 44.67773 | -1.09645; 44.65625 | 56 | 3856 | Meso | 10.8-20.8 |
Thau | 3.53828; 43.35232 | 3.57339; 43.3637 | 1295 | 125 | Micro | 8.6-24.9 |
*seasonal mean
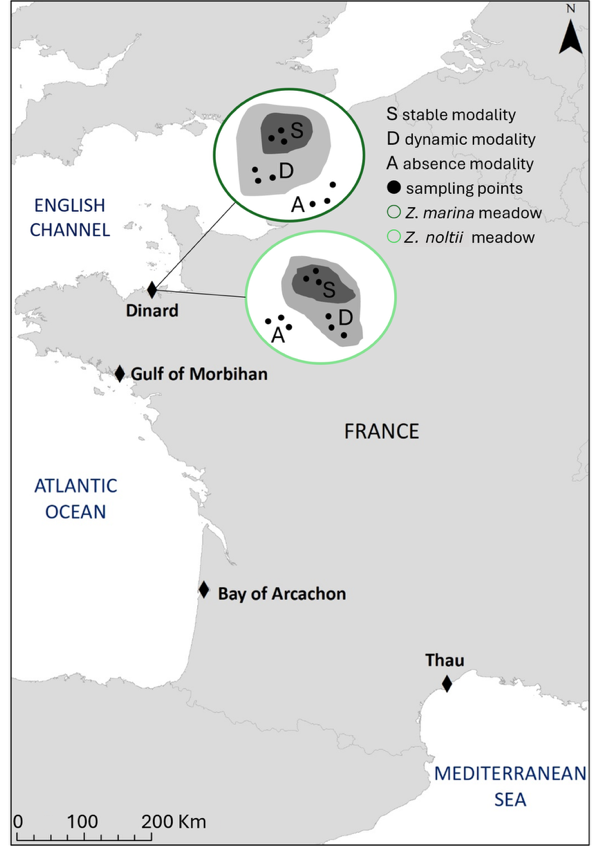
Figure 1 - Location of the four study sites in France. Nine flux measurements were carried out at each site in each habitat (Z. marina and Z. noltii), in three modalities: (S) stable and (D) dynamic parts of the seagrass meadow and (A) adjacent bare area. Modality D could not be identified in Arcachon.
Sediment-water fluxes measurement
Fluxes were measured during immersion using benthic chambers (0.13 m2 surface area) during photosynthetic (transparent dome – light incubation) and respiratory (opaque dome - dark incubation) periods successively. The incubation time varied between 1.5 and 2.5 hours depending on the site and the season. Salinity was continuously recorded during light and dark incubations using a AQUALABO-C4E sensor. Oxygen and temperature were measured continuously (1 min frequency) within chambers using a HOBO ® U26-001 logger. O2 fluxes were calculated as the slope of the regression reported to incubation time and chamber surface area. Alkalinity and pH were measured at the beginning and the end of the incubation to calculate DIC fluxes as the difference in concentration, relative to incubation time and chamber surface area. pH was measured on board using a multi-parameter logger (WTW® Multi 3620 IDS) equipped with a pH sensor (WTW® IDS SensoLyt 900-P). Alkalinity (mol.kg-1) was measured by pH-metric titration using the Titroline 7000 (SI Analytics, Germany), according to Dickson et al. (2007). DIC was obtained from total alkalinity and pH using conversion from the seacarb package (Gattuso et al., 2024). It should be noted that this method is based on the assumption that no distinction is made between DIC production from respiration processes and DIC production or consumption associated with carbonate dissolution or precipitation. Oxygen and DIC fluxes measured in dark conditions were expressed as community respiration (CR) whereas fluxes in light conditions corresponded to the net community production (NCP). Gross community production (GCP) was obtained by subtracting the CR from the NCP. Underwater photosynthetic photon flux density (PPFD, µmol m-2 s-1) was measured continuously in the chambers during the light incubation period using a spherical underwater quantum sensor (LI-COR193).
Habitat characterization
Habitat was characterized though the assessment of seagrass traits (shoot density, percentage cover, above and below ground dry biomass and leaf area index), macroalgae dry biomass, organic matter content in the sediment (loss on ignition method: 6h at 520°C), as well as density and ash free dry mass (AFDM) of infauna collected in benthic chambers at the end of each incubation series. Macrofaunal samples were collected with a small corer (0.05 m2) in the top 10 cm of the sediment. Organisms were sieved on 1 mm and identified to the species level. AFDM was determined for the whole sample after burning 5h at 450°C. All detailed protocols can be found in Lacoste et al., (2023). The results and their description, including the raw data and graphics, can be found in companion papers (Lacoste et al., 2023, 2024, 2025), and are not therefore described in detail in this article (but see Table 2 for partial results). The purpose of the data in the present study is to aid the interpretation of the variability of the measured fluxes.
Table 2 - Min and max values of organic matter in the sediment (OM %), shoot density (shoots per m2) and macrofauna biomass (AFDM: ash free dry mass in g) measured in the meadows at each site independently of season and modality (S and D).
Site | OM | Shoot density | Macrofauna AFDM | |||
|---|---|---|---|---|---|---|
Z. marina | Z. noltii | Z. marina | Z. noltii | Z. marina | Z. noltii | |
Dinard | 1.5-18.7 | 0.7-4.2 | 31-1075 | 157-8800 | 0-208 | 3-1044 |
Gulf of Morbihan | 2.6-29.2 | 2.1-8.2 | 0-262 | 198-10742 | 0-25 | 8-340 |
Arcachon | 4.9-11.8 | 4.1-22.4 | 0-81 | 198-10544 | 3-55 | 0-624 |
Thau | 3.6-7.6 | 2-6.4 | 0-225 | 199-4974 | 0-203 | 0-163 |
Data analysis
Statistical analyses were performed using R v4.3.1 (R core team 2023). Statistical differences in CR and GCP between sites and modalities were examined using linear mixed effect models with the package lme4 (Bates et al., 2015), with Site and Modality as crossed fixed effects and Season as a random effect. The effect of factors was considered significant when the null vs main effect models (using maximum likelihood estimation) differed significantly (ANOVA p < 0.05). Where the effect of interaction was significant, the least-squares means were calculated using the emmeans package (Lenth, 2023) to test for inter- and intra-site differences for each modality. The goodness of fit of the model was checked by examining the residual component (Quinn and Keough, 2002).
The relationships between benthic metabolism (CR & GCP) and habitat characteristics and environmental factors (independently of site and season) were explored separately for seagrass (S and D modalities) and bare sediment (A modality). We employed multiple linear models with the stepwise selection procedure (backward direction) of the MASS package (Venables and Ripley, 2002), to select the best model based on the Akaike Information Criterion (AIC) (Akaike, 1973). Variables without a direct apparent effect on fluxes (e.g. no direct effect of macrofauna on GCP) were discarded from the model. Absence of multicollinearity of predictor variables was verified using variance inflation factors (final VIFs < 3).
Analyses were carried out on the basis of O2 fluxes only (not DIC), given the continuous nature of O2 measurement and the higher precision in the data. Complete results for three seasons (Summer and Autumn 2020 and Spring 2021 for all sites) were used. All considered GCP fluxes were measured at PPFD values (Table 3) at or above photosynthetic saturation intensities for Z. marina and Z. noltii according to the literature (Plus et al., 2005; Léger-Daigle et al., 2022). Light was therefore not considered in the analysis assuming that near maximal community fluxes were recorded.
Table 3 - Mean underwater Photosynthetic Photon Flux Density (PPFD in µmol m-2 s-1) measured during flux experiments at each site and each season in Z. marina and Z. noltii habitats.
Site | Season | Z. marina | Z. noltii |
|---|---|---|---|
Dinard | 2020 - Summer | 331.9±25.2 | 404.6±219.9 |
2020 - Autumn | 296.2±48.6 | 257.5±67.2 | |
2021 - Spring | 343.4±21.8 | 715.9±37.5 | |
Gulf of Morbihan | 2020 - Summer | 777.1±173.1 | 1011.6±79.8 |
2020 - Autumn | 509.9±54.3 | 527.8±31.5 | |
2021 - Spring | 506.3±35.9 | 1369.6±136.1 | |
Arcachon | 2020 - Summer | 585.3±24.3 | 944.9±255.5 |
2020 - Autumn | 182.1±0.4 | 684.4±371.4 | |
2021 - Spring | 593.3±15.4 | 614.0±213.4 | |
Thau | 2020 - Summer | 371.3±70.5 | 743.6±108.5 |
2020 - Autumn | 280.0±30.4 | 100.2±4.5 | |
2021 - Spring | 358.2±59.4 | 405.1±84.6 |
In seagrass habitat, the community respiration (CRQ) and the community photosynthesis quotients (CPQ) were calculated as the standardised major axis (Warton et al., 2006) of the regression between CRDIC and CRO2 and between GCPO2 and GCPDIC respectively, using the smatr package (Warton et al., 2012).
Results
Common seasonal trend across sites
Diel variations of metabolism were observed for all habitats, with O2 consumption in the dark and O2 production during the light incubations (Figures 2, 3). For both Z. marina and Z. noltii, the main pattern of fluxes for all sites and all modalities consisted of higher values during the summer period, corresponding to the highest water temperature. Maximum values of |CR| were 16.39 mmol O2 m-2 h-1 for Z. marina and 13.03 mmol O2 m-2 h-1 for Z. noltii, observed during summer in Thau and Arcachon respectively. Maximum values of GCP were 23.24 mmol O2 m-2 h-1 for Z. marina and 28.39 mmol O2 m-2 h-1 for Z. noltii observed during summer in Gulf of Morbihan and Arcachon respectively. The lowest values of fluxes were generally observed during the winter, even though seasonal differences are less evident for the GCP as compared to the CR (Figures 2, 3).
In the presence of seagrass (Stable and Dynamic modalities) GCP values were always higher than CR (with the sole exception of Dinard during the summer). In the absence of seagrass (A modality), CR > GCP was observed several times (about 40% of the total fluxes), indicating that the measured heterotrophic processes were more important than the autotrophic process at the time of measurement.
Spatial variability of sediment-water fluxes
The Site × Modality interaction was significant for both the CR and GCP fluxes in the Z. marina and Z. noltii habitats (Table 4). Within Z. marina habitats, CR values were overall highest in Thau regardless of the modality (Figure 2, Table S2), and GCP values were lower in Dinard and Arcachon compared to Gulf of Morbihan and Thau (Figure 2, Table S4). For Z. noltii GCP and CR were significantly lower in Dinard compared to Thau and Gulf of Morbihan (Figure 3, Tables S3, S5).
Table 4 - Results of the linear mixed effect models testing the effect of Site (n=4), Modality (n=3) and their interaction on community respiration (CR) and gross community production (GCP) during immersion in Z. marina and Z. noltii habitats. Df: degrees of freedom, Chisq=chi-square value. See Tables S2-S5 for pair-wise comparisons.
Species | Factor | CR | GCP | ||||
Df | Chisq | p-value | Df | Chisq | p-value | ||
Z. marina | Site x Modality | 13 | 44.018 | <0.001 | 13 | 71.56 | <0.001 |
Z. noltii | Site x Modality | 13 | 69.575 | <0.001 | 13 | 76.431 | <0.001 |
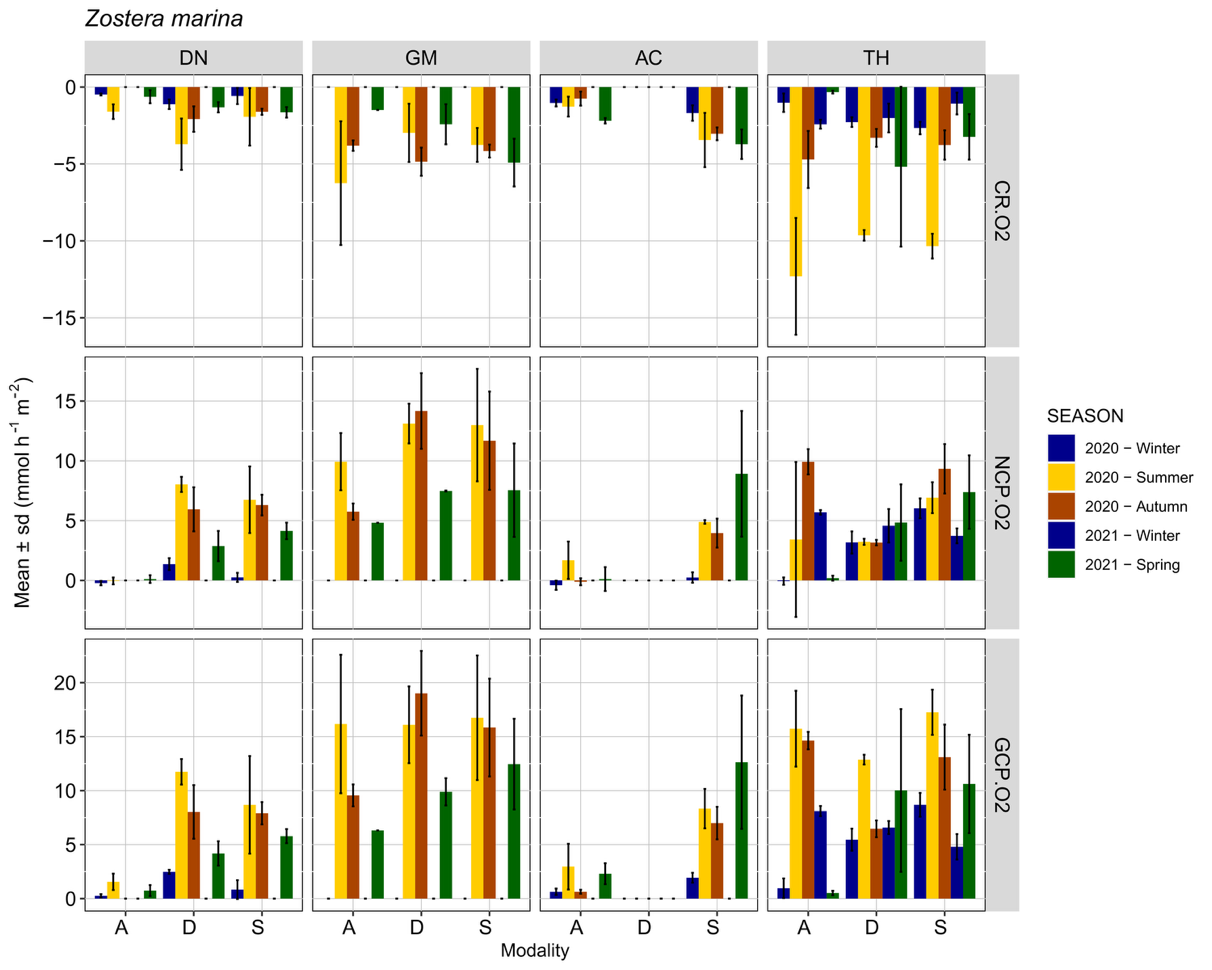
Figure 2 - Mean ± sd (n=3) community respiration (CR.O2), net community production (NCP.O2) and gross community production (GCP.O2) measured with the O2 method for Z. marina during different seasons for three modalities: A: bare sediment, D: dynamic and S: stable, in Dinard (DN), Gulf of Morbihan (GM), Arcachon (AC) and Thau (TH).
Mean CR in Z. marina was not significantly affected by the within-habitat position (Table S2). This absence of difference is likely due to important CR observed at modality A in Gulf of Morbihan and Thau. In Z. noltii, CR was lower in absence of seagrass (A modality), with no significant difference in the Gulf of Morbihan (Table S3). In Dinard, there was also an effect of the within-habitat position (Z. noltii), with higher CR in modality S compared to modality D (almost doubled in summer, Figure 3, Table S3).
Mean GCP was significantly lower in modality A of each species compared to the other two modalities (Figure 2,3, Table S4), except in Thau for Z. marina where an important biomass of macroalgae was observed in the bare sediment area in summer and winter (>300 mgDW.m-2). Position within the seagrass bed (modality S vs. D) did not significantly influence GCP either for Z. marina or Z. noltii (Tables S4, S5), although large variations, by as much as double the value, were sometimes observed (Figure 3).
With some exceptions, standard deviation was low (Figures 2, 3), illustrating a good measurement repeatability through use of the benthic chamber method.
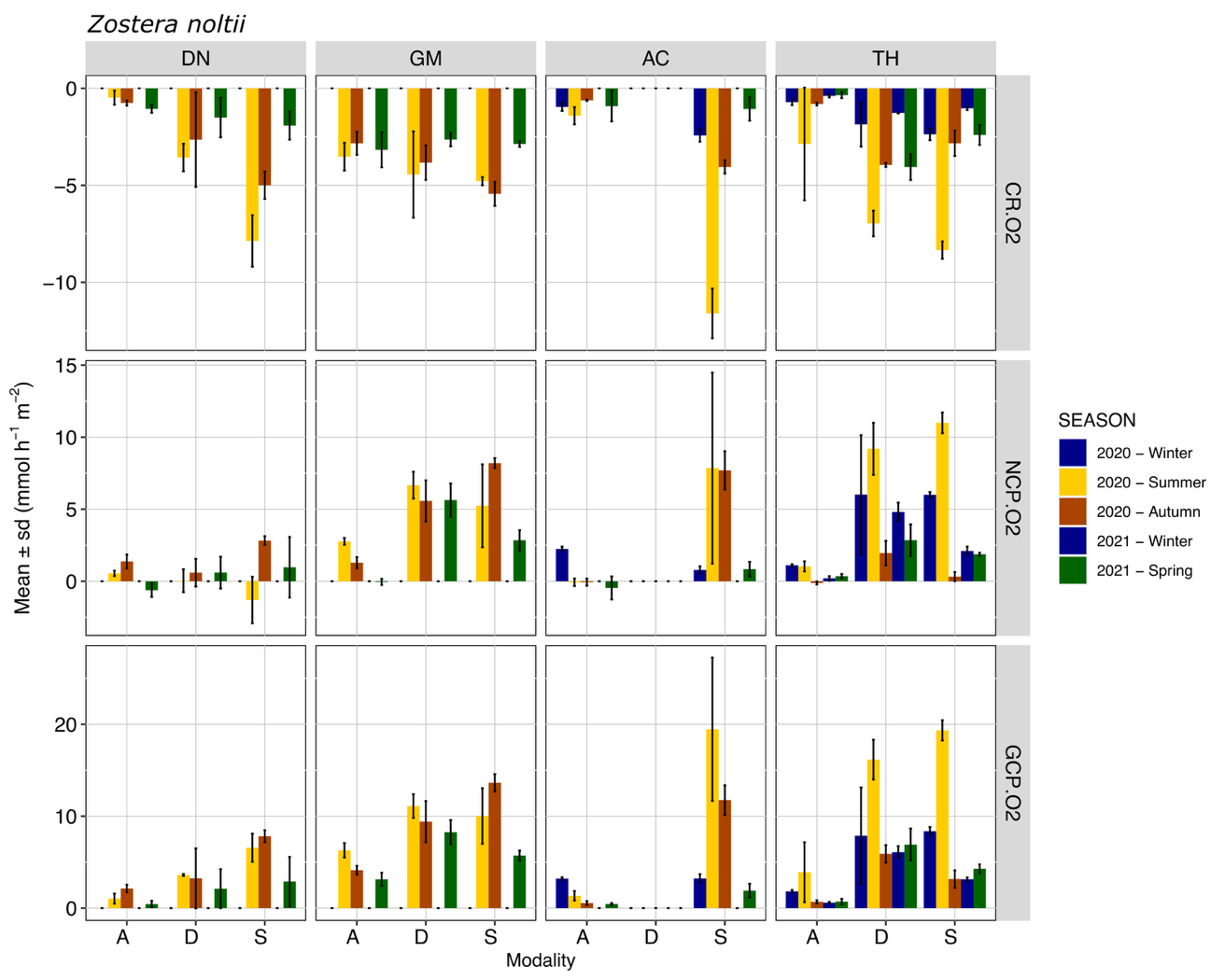
Figure 3 - Mean ± sd (n=3) community respiration (CR.O2), net community production (NCP.O2) and gross community production (GCP.O2) measured with the O2 method for Z. noltii during different seasons and for three modalities: A: bare sediment, D: dynamic and S: stable, in Dinard (DN), Gulf of Morbihan (GM), Arcachon (AC) and Thau (TH).
Influence of habitat on benthic metabolism at the sediment water interface
Explanatory variables of O2 fluxes
Overall, water temperature was the most consistent significant factor across habitats and species (Table 5), explaining both CR and GCP. Other factors such as salinity, percentage of organic matter in the sediment and algal biomass showed variable significance depending on habitat and species (Table 5). Macrofauna (density) only had a significant effect on CR in Z. marina seagrass habitat (Table 5). Leaf biomass for both Z. marina and Z. noltii and leaf area index for Z. noltii were the two significant morphometric trait descriptors of GCP in seagrass habitats (Table 5).
Table 5 - Significant results of the multiple regressions testing the effect of measured environmental parameters on community respiration (CR) and gross community production (GCP) in Z. marina and Z. noltii habitats. Habitat: seagrass = grouped dynamic and stable modalities (S, D), bare = modality A. Only significant models (p-value < 0.05) are presented (i.e. Z. noltii GCP in bare sediment was not significant). Initial models, the relative importance of each predictor in the final models and number of samples (n) are reported in Table S1. F: Fisher statistic, RSE: residual standard error, adj. R2: adjusted R2.
Species | Habitat | Model terms | Estimate | Std. Error | p-value |
Z. marina | seagrass | 1. CR: F=12.74; RSE= 1.19; adj. R2=0.74 | |||
Water temperature | 2.78 × 10-1 | 5.34 × 10-2 | < 0.001 | ||
Salinity | 3.01 × 10-1 | 9.24 × 10-2 | < 0.005 | ||
Leaf biomass | 9.22 × 10-3 | 5.77 × 10-3 | ns | ||
Root biomass | 2.87 × 10-3 | 2.27 × 10-3 | ns | ||
% organic matter | 1.15 × 10-1 | 4.58 × 10-2 | < 0.05 | ||
Infauna density | -3.19 × 10-4 | 1.16 × 10-4 | < 0.05 | ||
Z. marina | bare | 2. CR: F=13.93; RSE=1.62; adj. R2=0.73 | |||
Water temperature | 3.20 × 10-1 | 8.01 × 10-2 | < 0.005 | ||
Algal biomass | 1.47 × 10-2 | 3.46 × 10-3 | < 0.005 | ||
% organic matter | 2.16 × 10-1 | 1.31 × 10-1 | ns | ||
Z. marina | seagrass | 3. GCP: F=7.81; RSE=3.42; adj. R2=0.52 | |||
Water temperature | 0.38 × 10-1 | 1.45 × 10-1 | < 0.05 | ||
Salinity | 0.38 × 10-1 | 2.54 × 10-1 | ns | ||
Shoot density | -5.74 × 10-3 | 3.00 × 10-3 | ns | ||
Leaf biomass | 3.40 × 10-2 | 1.44 × 10-2 | < 0.05 | ||
Z. marina | bare | 4. GCP: F=4.941; RSE=4.785; adj. R2=0.36 | |||
Water temperature | 4.30 × 10-1 | 2.33 × 10-1 | ns | ||
Salinity | 9.31 × 10-1 | 4.16 × 10-1 | < 0.05 | ||
Z. noltii | seagrass | 5. CR: F=19.15; RSE= 1.091; adj. R2=0.83 | |||
Water temperature | 2.89 × 10-1 | 4.40 × 10-2 | < 0.001 | ||
Leaf Area Index | 2.03 × 100 | 5.58 × 10-1 | < 0.005 | ||
Algal biomass | -8.18 × 10-2 | 2.17 × 10-2 | < 0.005 | ||
% organic matter | -8.58 × 10-2 | 5.45 × 10-2 | ns | ||
Infauna density | -8.69 × 10-5 | 5.02 × 10-5 | ns | ||
Infauna AFDM | 2.17 × 10-2 | 1.18 × 10-2 | ns | ||
Z. noltii | bare | 6. CR: F=4.81; RSE=0.83; adj. R2=0.47 | |||
Water temperature | 1.39 × 10-1 | 4.85 × 10-2 | < 0.05 | ||
Infauna density | 1.85 × 10-4 | 9.03 × 10-5 | ns | ||
Infauna AFDM | 4.08 × 10-2 | 1.99 × 10-2 | ns | ||
Z. noltii | seagrass | 7. GCP: F=12.9; RSE=2.92; adj. R2=0.67 | |||
Water temperature | 0.38 × 10-1 | 4.56 × 10-2 | < 0.01 | ||
Leaf Area Index | 34.6 × 10-1 | 14.8 × 10-1 | < 0.05 | ||
Algal biomass | -1.22 × 10-1 | 5.57 × 10-2 | < 0.05 | ||
Leaf biomass | 4.44 × 10-2 | 1.75 × 10-2 | < 0.05 | ||
Comparison of O2 and DIC fluxes measurement: CR and CP quotients
Considering results from all seasons (Figure 4) the community respiration quotient (CRQ) varied between 0.73 and 1.07 in Z. marina habitat and between 1.12 and 1.66 in Z. noltii habitat (Table 6) with the slope significantly different from 1 in Z. noltii. The regression slopes between GCPDIC and GCPO2 (CPQ) were different from 1 for both species with values between 1.01 and 1.41 for Z. marina and between 1.01 and 1.60 for Z. noltii. Except for the CRQ in Z. marina, the deviation of the quotients from 1 indicated an imbalance between production and consumption processes.
For both species the CRQ was lower during spring and summer (Table 6). The CPQ was lower in autumn for Z. marina and in summer for Z. noltii (Table 6). When considering each season separately, most of the regressions were not significant.
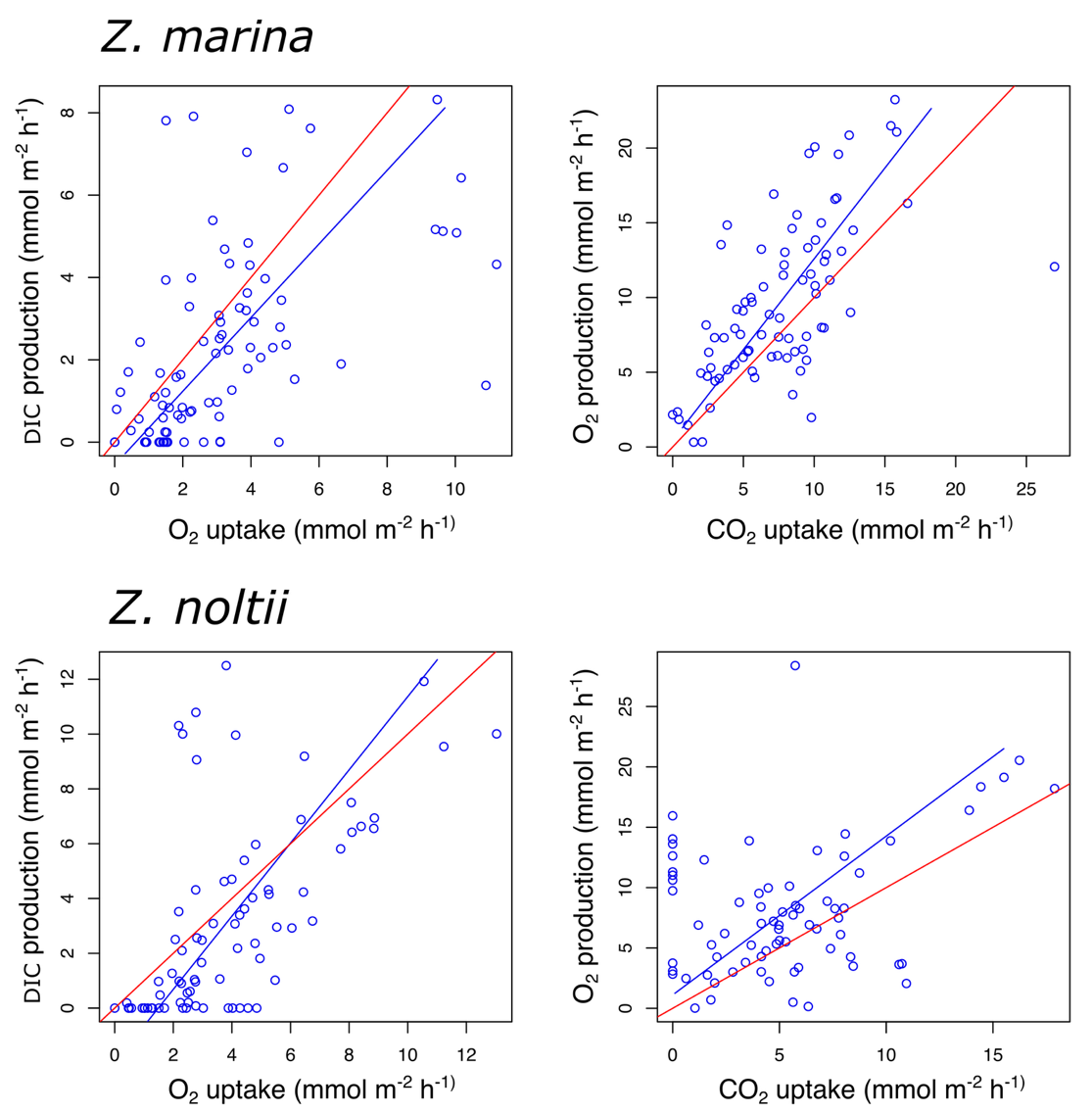
Figure 4 - Relationships between DIC production and O2 uptake during dark (CRQ, left panels), and O2 production and DIC uptake (CPQ, right panels) during light incubations for Z. marina (top panels) and Z. noltii (bottom panels). The blue line is the linear regression fitted on the data. The red line corresponds to y=x.
Table 6 - Estimation of the community respiration (CRQ) and community photosynthesis (CPQ) quotients from the standardized major axis using results from all four sites for grouped dynamic and stable modalities (S, D), per season, and for all seasons.
Species | Season | N | Slope | Lower limit | Upper limit | R2 | p | |
CRQ | Z. marina | 2020 Winter | 15 | 1.15 | 0.67 | 1.97 | 0.10 | 0.24 |
2020 Summer | 21 | 0.78 | 0.51 | 1.20 | 0.14 | 0.09 | ||
2020 Autumn | 21 | 1.49 | 1.04 | 2.15 | 0.40 | 0.002 | ||
2021 Spring | 21 | 0.63 | 0.40 | 0.98 | 0.08 | 0.20 | ||
All seasons | 78 | 0.88 | 0.73 | 1.07 | 0.28 | <0.001 | ||
Z. noltii | 2020 Winter | 9 | 6.76* | 3.09 | 14.77 | 0.06 | 0.50 | |
2020 Summer | 21 | 1.33 | 0.86 | 2.05 | 0.13 | 0.11 | ||
2020 Autumn | 21 | 1.49 | 0.99 | 2.25 | 0.24 | 0.02 | ||
2021 Spring | 20 | 1.32 | 0.94 | 1.84 | 0.53 | <0.001 | ||
All seasons | 71 | 1.36 | 1.12 | 1.66 | 0.34 | <0.001 | ||
CPQ | Z. marina | 2020 Winter | 13 | 1.33 | 0.99 | 1.78 | 0.80 | <0.001 |
2020 Summer | 21 | 1.39 | 1.01 | 1.91 | 0.55 | <0.001 | ||
2020 Autumn | 21 | 0.88 | 0.62 | 1.26 | 0.44 | 0.001 | ||
2021 Spring | 20 | 1.29 | 0.86 | 1.96 | 0.26 | 0.02 | ||
All seasons | 75 | 1.19 | 1.01 | 1.41 | 0.46 | <0.001 | ||
Z. noltii | 2020 Winter | 9 | 1.13 | 0.52 | 2.46 | 0.08 | 0.46 | |
2020 Summer | 21 | 1.11 | 0.71 | 1.70 | 0.11 | 0.14 | ||
2020 Autumn | 20 | 1.43 | 0.91 | 2.24 | 0.12 | 0.14 | ||
2021 Spring | 17 | 1.15 | 0.68 | 1.94 | 0.005 | 0.79 | ||
All seasons | 67 | 1.27 | 1.01 | 1.60 | 0.13 | 0.003 |
*Outlying value excluded from mean calculation
Discussion
Seagrass habitats play critical ecological roles in several ecosystem processes such as oxygen production via photosynthesis and O₂ release into the water, and carbon cycling regulation via DIC capture through photosynthesis, storing it in their biomass and sediments. The benthic chamber method presented here allows the estimation of instantaneous sediment-water exchanges at the community-level and has been widely used in different habitats and environments (e.g. Smith et al., 2009; Ouisse et al., 2014; Grenz et al., 2021). Using this methodology, we provided an extensive dataset of benthic O2 and DIC fluxes for two emblematic seagrass species, Z. marina and Z. noltii, covering a wide range of environmental conditions. The strength of this study lies in the use of a standardized, common method deployed across four sites spread across a broad latitudinal gradient and over different seasons, allowing global analyses without bias.
Benthic metabolism at immersion in seagrass habitat
Table 7 - Range of community respiration (CR) and gross community production (GCP) values measured using the immersed benthic chambers method and O2 measurements throughout the distribution area of Z. marina and Z. noltii over different seasons in existing literature.
Study | Habitat | Area | |CR| (mmol O2 m-2 h-1) | GCP (mmol O2 m-2 h-1) |
This study | Z. marina | Multi sites (Atlantic & Thau lagoon) | 0.05 to 16.4 | 0.16 to 23.2 |
Z. noltii | 0.2 to 13.1 | 0.02 to 28.4 | ||
Z. noltii | Western English Channel | 0.5 to 15 | 0.5 to 18 | |
Z. marina | Bay of Brest | 2.5 to 5 | 3 to 13 | |
Z. marina | Bages lagoon | 5 to 10 | 15 to 30 | |
Z. noltii | Ayrolle lagoon | 2.7 to 10 | 15 to 30 | |
Z. noltii | Thau lagoon | 1.6 to 15.1 | 3.1 to 31.3 | |
Undetermined eelgrass | Baltic sea | 0.1 to 3.8* | 1.7 to 2.7 | |
Z. marina | Gulf of Finland | 1.4 to 5.9 | 3.7 to 8.5 |
*Graphical estimation
The community respiration (CR) and gross community production (GCP) values measured during this study are within the range of previous values obtained in temperate regions for these two species using the same method (Table 7). As observed in these studies, the highest values were obtained most of the time during the warmest periods, confirmed by the strong positive correlation between water temperature and fluxes. This is consistent with the general pattern of seagrass metabolism, which shows higher gross primary production during the summer months in both tropical and temperate ecosystems (Ward et al., 2022).
Overall, the benthic metabolic activity was enhanced in the presence of Z. marina and Z. noltii compared to adjacent uncolonized sediments. Exceptions were observed in the Thau lagoon and the Gulf of Morbihan, where the fluxes were similar in the presence or absence of seagrass. This was mainly due to the presence of macroalgae, which may play a similar role to seagrass in increasing habitat complexity. This confirms the important role of seagrass (and macroalgae) as an ecosystem engineer - sensu Jones et al., (1994), i.e. resource modifiers for other associated species - and their fundamental role in carbon and oxygen cycle (Fourqurean et al., 2012). This investigation brings additional confirmation that conservation efforts aimed at protecting and restoring seagrass habitats are essential for preserving the valuable ecosystem functions and services they provide. These results also highlight the need for habitat-specific measurements of benthic fluxes, often scarce in the literature to properly quantify habitat ecosystem functions. Together with habitat mapping, such information would provide a unique opportunity to capture the global functioning of coastal ecosystems.
Given the variability in morphological and biological traits of Z. marina between the four sites (Lacoste et al., 2025), one might have expected very different functional responses in terms of O2 and DIC metabolism. On one hand, the morphological characteristics of Z. marina in Dinard have been described as adapted to stressful conditions (i.e. strong hydrodynamic conditions), with important shoot density and below-ground biomass for anchoring, with limited leaf surface area (see also Boyé et al., 2022). Conversely, the seagrass population that developed in the more turbid and calm conditions of Arcachon showed low shoot density but long and wide leaves, thereby maximizing their access to light. These different - and at the opposite of the ecological spectrum - strategies, expressed by different phenotypes, were not reflected in the ecological functioning at the community-level, since no significant difference was observed in the measured fluxes at immersion between these two sites. Similarly, the phenotypic difference observed for Z. marina at the modality scale in Dinard, with greater shoot density in the dynamic as opposed to the stable area, was not observed for fluxes. This is a sign of high phenotypic plasticity of this species, able to adapt to local conditions without altering its capacity to efficiently use resources. Seagrasses are however engineer species that host a highly diverse species assemblage, that include other primary producers such as epiphytes or microphytobenthos, as well as heterotrophic organisms, from bacteria to epifauna. Thus, GCP and CR derive from complex processes operated by different interacting organisms, and are also influenced by the environment (Rodil et al., 2021) such that seagrass characteristics alone may not be good proxies of ecosystem functioning.
Predicting flux variability at the global scale
In most cases, benthic metabolism differed markedly between vegetated areas and adjacent sediments. As such, we explored correlations between environment and fluxes separately within the meadow (modalities S and D), and in the bare area (modality A). Models in modality A were less well fitted than in seagrass, except for community respiration (CR) near Z. marina (R2: 0.74). The presence of macroalgae in Thau appears to have driven this correlation by increasing habitat complexity and possibly establishing heterotrophic communities (Friedrich, 2012 and references therein). Considering the regression analysis, the role of macrofauna does not appear to be the most important in our study, so other communities may be more important. The positive role of temperature on CR in this modality suggests an important role of meiofauna and bacteria inhabiting sediments and macroalgae, whose activity could be increased in warmer conditions (Rivkin and Legendre, 2001; Del Giorgio et al., 2006). Conversely the poor prediction of gross community production (GCP) in bare sediments may have suffered from the lack of quantification of primary producers such as microphytobenthos, which could be an important contributor to photosynthesis in coastal areas (Bartoli et al., 2003; Spilmont et al., 2006).
Of the total observed variance in benthic metabolic activity in seagrass, 52% to 83% was explained by the factors considered in our measurements, the most important being temperature, leaf biomass and surface area. This supports the direct effect of seagrass biomass on benthic primary production and respiration, as previously shown in studies that have compared vegetated and nearby unvegetated sediments (Stutes et al., 2007; Hume et al., 2011; Delgard et al., 2016). The positive effect of temperature on GCP is likely due to both increased photosynthesis in warmer water and to co-correlation, with seasonally-driven morphometric seagrass characteristics, in particular leaf growth. It should be noted that measurements were likely carried out during the optimal temperature range for these species (Berger et al., 2020), with no apparent stress during the studied period (Lacoste et al., 2025). Photosynthetic capacity is however known to start declining above a certain temperature, together with plant biomass loss (Lee et al., 2007; Collier et al., 2017). We therefore stress the importance of considering the evolution of seagrass benthic metabolism and their role in benthic-pelagic coupling with respect to the increased sea water temperatures predicted under climate change. This is particularly true at the distribution range edge of the species, such as in the Thau lagoon for Z. marina.
Finally, given the lack of prediction of GCP variation in our model (R2: 0.52 and 0.67 for Z. marina and Z. noltii, respectively), one might expect to improve this predictability by quantifying other primary producers that may contribute significantly to coastal primary production, such as epiphytes and microphytobenthos, as mentioned above. It was for example previously found that epiphytic biofilm developing on seagrass leaves have an effect on photosynthesis and respiration rate (Hansen et al., 2022) but that both bare leaves and epiphytes might receive different amounts and nature of light, and thus respond differently to environmental changes (Brush and Nixon, 2002). Despite this limitation, our dataset provides linear regression models that can be used to make predictions about benthic metabolism in seagrass habitats under a given set of environmental conditions with knowledge of some seagrass morphometric traits.
The oxygen-carbon balance
Most existing studies of metabolic processes of macrophyte communities rely on measuring O2 consumption and production rather than carbon (e.g. Hansen et al., 2000; Santos et al., 2004; Barrón et al., 2006), mainly due to the relatively straightforward measurement method of the former. The close correlation between DIC and O2 release or demand due to respiration and photosynthesis at the water-sediment interface suggests that DIC and O2 fluxes in sediments are closely coupled processes. As we are often more interested in carbon cycling, CO2 uptake and release are therefore typically derived from O2/CO2 conversion, with the community photosynthesis quotient (CPQ) and community respiration quotient (CRQ) factors set to 1 which is the theoretical value expected. However, there are challenges in using this conversion factor when autotrophic and heterotrophic organisms coexist – such as in seagrass habitats – and for which situations the CRQ values tend to deviate from 1 (Trentman et al., 2023). For example, anaerobic reduction of sulphate may underestimate respiratory losses while overestimating net production. As part of this study, we have jointly estimated O2 and DIC fluxes, providing a reliable estimate of CRQ and CPQ ratios across a large gradient of ecosystem variations spanning most of the Z. marina and Z. noltii range in Europe.
The measured values of both CRQ and CPQ were within the range of values reported in the literature for seagrasses (Martin et al., 2005; Ouisse et al., 2014; Le Fur, 2018), deviating from 1. For all seasons combined, CPQ was > 1 for both species, indicative of higher O2 production compared with DIC consumption. Such an imbalance may be due to several processes such as nitrate reduction processes releasing O2 during biosynthesis associated with primary production (Trentman et al., 2023), or DIC consumption partly compensated by benthic alkalinity fluxes due to anaerobic processes. The CRQ was > 1 for Z. noltii, indicating the possibility of organic matter decomposition using elements other than O2 as terminal electron acceptors, such as nitrate or sulphate. This was also observed in winter and autumn for Z. marina, indicative of a potential increase of anaerobic processes during these periods. This hypothesis could be supported by the fact that seagrass meadows are known to be sites of high organic matter remineralisation, trapping and concentrating organic particles (Duarte et al., 2005).
Our results show that measuring O2 fluxes only and assuming a CPQ of 1 would have led to overestimating community carbon assimilation in seagrass. For a better comprehension of community metabolism, it is therefore advised to measure O2 and DIC fluxes simultaneously such that the ratio could inform on the biogeochemical processes mentioned above. Although our summary table (Table 6) provides a range of values for CRQ and CPQ at the scale of seagrasses distribution in France, predicting when and where quotients may vary as a function of environmental conditions requires further comparison of O2 and DIC measurements at high sampling rates.
Conclusion
Seagrasses are key species in the functioning of coastal habitats, with their importance in carbon sequestration and ocean acidification mitigation now fully recognized. Accounting for spatial and temporal variability in metabolism is therefore particularly important for managers, given the growing interest in seagrass restoration and innovations towards nature-based solutions to mitigate ocean acidification and sequester carbon in the sediments. This study provides some clues and evidences to better understand whether gross community production and respiration vary predictably over time and at different spatial scales. To the best of our knowledge, this study is the first to deploy standardised in situ measurements, while factoring in seasonal effect at different sites and habitats, varying both in terms of macrophyte cover (seagrass vs bare) and meadow heterogeneity (stable vs dynamic area seagrass bed areas). At the global scale, some of the variability remains unexplained, probably due to the lack of direct quantification of certain community components, such as the microphytobenthos or the bacterial compartment. Functional diversity of macrofauna could also help explain part of the variability (Hillman et al., 2021). Site-specific models could actually increase the predictability of fluxes, but in most situations, data are scarce. The models proposed here therefore represent an imperfect alternative, which will nonetheless provide a better understanding of the role of seagrass beds in the functioning of coastal ecosystems. We also provide a reliable range of values with associated errors for converting O2 to DIC when estimating seagrass carbon budgets from O2 measurements. Ongoing global changes in environmental conditions and the extent of seagrass meadows will undoubtedly require additional efforts to estimate benthic metabolism as a function of environmental conditions and increasing water temperature.
Acknowledgement
This study is part of the Life Marha project funded by the European Union. The authors thank Thomas Ribeiro dos Santos for his help in weighing the macrofauna, Céline Cordier for preparing multiple fieldwork expeditions, and Lionel Alano for his help and boat in the Morbihan Gulf.
Preprint version 4 of this article has been peer-reviewed and recommended by Peer Community In Ecology (https://doi.org/10.24072/pci.ecology.100755; Hubas, 2025).
Data, script and code
Data and supplementary information are available at https://doi.org/10.17882/97962 (Lacoste et al., 2023). Supplementary Tables and code are available at 10.5281/zenodo.13354064 (Lacoste, 2025, V4).
Funding
The authors declare that they have received no specific funding for this study.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.