
 CC-BY 4.0
CC-BY 4.0
Introduction
In bacteria, at the onset of chromosome replication initiation, replicative helicases are loaded onto the DnaA-oriC nucleoprotein platform with the assistance of helicase loaders, such as DnaC in Escherichia coli (Arias-Palomo et al., 2013) and DnaI in Bacillus subtilis (Velten et al., 2003). These latter are derived from mobile elements and have replaced, in a few bacterial orders, the ancestral and widespread unrelated replicative helicase loader gene dciA (Brézellec et al., 2016). Bacteria lacking both dnaC/dnaI and dciA are uncommon and generally exhibit small genomes or host-associated lifestyles (Brézellec et al., 2016). Nevertheless, Brézellec and collaborators showed that in two bacterial orders, Cellvibrionales and Oceanospirillales, the lambdoid phage genes λO and λP (the latter encoding the λ phage replicative helicase loader) could have substituted for dciA (Brézellec et al., 2017). In the same vein, in Vibrionales, dciA could also have been frequently replaced by two viral genes (Tominaga et al., 2024). To date, however, Helicobacter pylori is the only extensively studied bacterium that appears to lack the ancestral helicase loader gene dciA or phage gene counterparts (Blaine et al., 2023).
While it’s presumed that Helicobacter pylori has lost the ancestral replicative helicase loader gene dciA, the unique features of its replicative helicase (HpDnaB) point to a different scenario. HpDnaB exhibits profound differences from classical bacterial replicative helicases. First, HpDnaB differs from canonical DnaB by having a large two-helix insertion, termed HPI, in its C-terminal ATPase domain. This insertion spans 34 amino acid residues (residues 400–433) when compared to Escherichia coli DnaB (Stelter et al., 2012). Second, while classical bacterial replicative helicases display a hexameric architecture, HpDnaB forms head-to-head double hexamers, i.e., a dodecameric architecture (Stelter et al., 2012). This leads me to hypothesize that dciA is still present in the H. pylori genome, but that it has co-evolved with HpDnaB, obscuring its typical Pfam domain signature (DUF721/PF0528) upon which its identification relies.
Using genomic synteny, HHpred, and AlphaFold, I demonstrate here that dciA is present in the H. pylori genome and that it co-localizes with the uvrC gene (excinuclease ABC subunit C), see Figure 1. I then discuss the implications of this finding.
Materials and methods
Strains of Helicobacter pylori
A large number of Helicobacter pylori strains have been sequenced and annotated. In this study, I focused on strains 26695 (ATCC 700392) (Tomb et al., 1997) and G27 (Baltrus et al., 2009).
Softwares
I used the HHpred tool via its web server (https://toolkit.tuebingen.mpg.de/tools/hhpred). HHpred allows protein domain and structure prediction; it utilizes pairwise alignment of Hidden Markov models (HMMs) to perform sensitive protein sequence searching (Gabler et al., 2020).
I used AlphaFold’s results stored in the UniProt database. AlphaFold is an artificial intelligence (AI) program developed by DeepMind for protein structure prediction (Jumper et al., 2021).
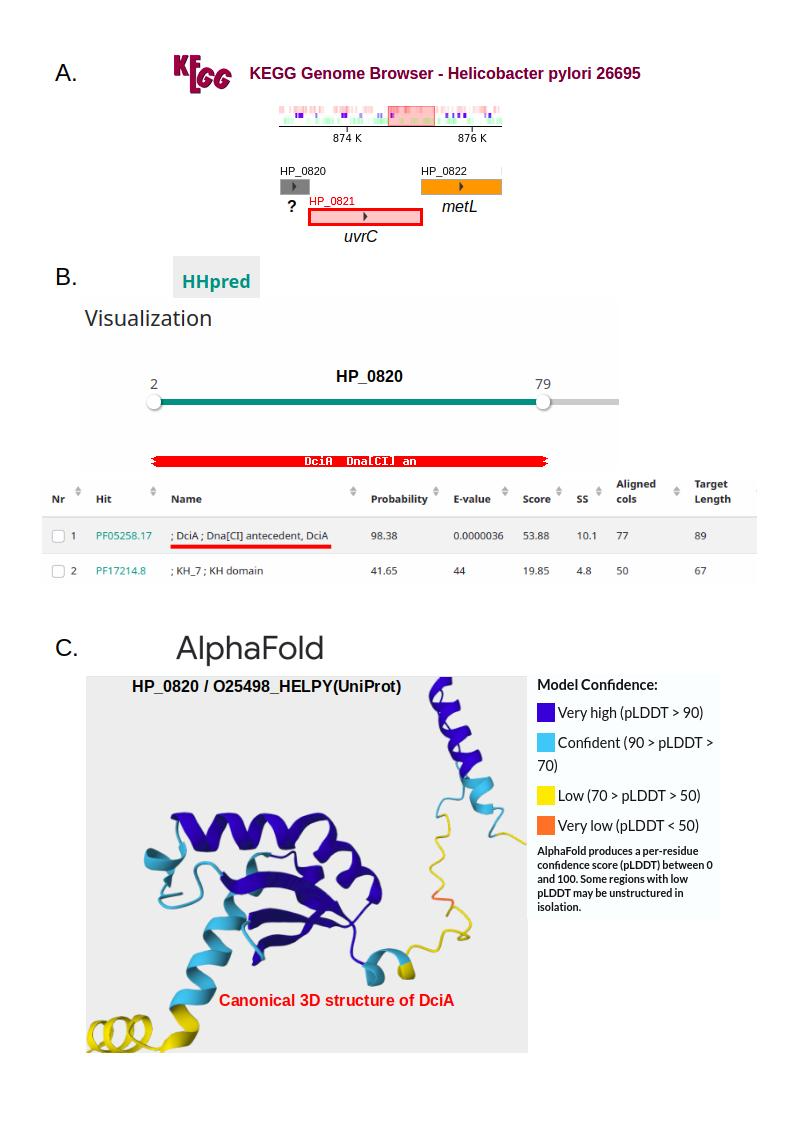
Figure 1 - It is presumed that Helicobacter pylori has lost the ancestral replicative helicase loader gene dciA. Using genomic synteny (see part A of the image), HHpred (see part B of the image), and AlphaFold (see part C of the image), I demonstrate that dciA is present in the H. pylori genome and that it co-localizes with the uvrC gene (excinuclease ABC subunit C). This finding significantly challenges the two current models of replication initiation in this bacterium. A. Many species in Campylobacterales, the order including H. pylori, show genomic co-localization of the dciA and uvrC genes. In H. pylori strain ATCC 700392 / 26695, uvrC (HP_0821) is surrounded by two genes: HP_0820, which encodes a protein annotated as ‘predicted coding region HP0820’, and HP_0822 (metL), encoding a protein annotated as homoserine dehydrogenase. B. I reannotated the protein encoded by HP_0820 using HHpred (with default parameters) against Pfam-A_37 (the latest Pfam release). HHpred analysis indicates that this protein contains the DciA-characteristic Pfam domain, i.e. PF05258/DUF721. This domain spans residues 2 to 79, with a probability of 0.986, indicating near-certain homology (see HHpred documentation). C. I focused on the AlphaFold-predicted structure of the protein encoded by HP_0820 (referred to as O25498_HELPY in UniProt) and observed the presence of the canonical DciA domain structure, which consists of one alpha helix, followed by two beta sheets, a third alpha helix, and a final beta sheet. Therefore, I concluded that HP_0820 is the dciA gene.
Results
Genomic co-localization of the dciA and uvrC genes is a feature of numerous Campylobacterales
To investigate the presence of dciA in the H. pylori genome, I began by examining its genomic context in dciA-hosting Campylobacterales species, the order to which H. pylori belongs. I aimed to leverage the genes frequently located near dciA to identify it in H. pylori. To achieve this, I utilized both the UniProt (UniProt Consortium, 2024) and KEGG (Kanehisa et al., 2024) databases, as detailed below.
To manage the substantial number of Campylobacterales proteomes, I concentrated on Campylobacterales ‘reference proteomes’ from UniProt. These proteomes offer a representative cross-section of taxonomic diversity and encompass model organisms and species of biomedical/biotech interest. UniProt contains 77 Campylobacterales reference proteomes, see Supplemental file 1 (query in ‘Proteomes’: (taxonomy_id:213849) AND (proteome_type:1), where ‘213849’ is the ‘Taxon ID’ for Campylobacterales and ‘1’ indicates ‘reference proteome’).
As shown in (Blain et al., 2022), DciA homologs across different phyla have low conservation in their primary amino acid sequences; consequently, tools such as BLASTP only retrieve very closely related homologs. DciA is therefore typically identified by its Pfam domain signature, DUF721/PF0528. Consequently, I searched the 77 previously identified reference proteomes for DciA-containing proteins using the following UniProt query: (taxonomy_id:213849) AND duf721 AND (keyword:KW-1185), where ‘KW-1185’ denotes ‘reference proteome’. UniProt returned 49 results, corresponding to 46 bacteria, see Supplemental file 2. Of these, 43 contained a single DciA protein, while 3 (Malaciobacter halophilus, Candidatus Campylobacter infans, and Helicobacter apodemus) contained two.
Table 1 - Each line represents a dciA gene. The ‘Organism’ column indicates the species to which dciA belongs. The ‘UniProt ID’ column refers to the UniProt ID of the considered dciA gene. The ‘Genomic context’ column indicates the KEGG annotation of the proteins expressed by the two genes located adjacent to dciA on the considered genome, with dciA referred to by its ordered locus name. As can be seen, the uvrC gene (in bold) is the most frequently located gene adjacent to dciA. Note that Malaciobacter halophilus and Candidatus Campylobacter infans host two dciA genes, while Helicobacter enhydrae expresses one DciA protein linked to two different KEGG loci.
Organism | dciA Ordered locus name | UniProt ID |
|---|---|---|
Aliarcobacter butzleri | Abu_0923 | A8ETA9 |
Arcobacter nitrofigilis | 508 | D5V0A6 |
Malaciobacter halophilus | 1117 | A0A2N1J241 A0A2N1J4B3 |
Poseidonibacter parvus | 1683 | A0A1P8KND2 |
Campylobacter iguaniorum | 669 | A0A076FH25 |
Candidatus Campylobacter infans | 1960 | A0A7H9CJ49 A0A7H9CKE6 |
Campylobacter curvus | 1995 | A7GX63 |
Campylobacter jejuni | 284 | Q0P909 |
Campylobacter lari | 284 | B9KFL2 |
Helicobacter enhydrae | A0A1B1U6Z8 A0A1B1U6Z8 | |
Helicobacter bizzozeronii | F8KQE8 | |
Helicobacter hepaticus | Q7VK74 | |
Wolinella succinogenes | Q7MR76 | |
Hydrogenimonas sp | A0A3G9GU81 | |
Sulfurimonas lithotrophica | A0A5P8P2C0 | |
Sulfurimonas marina | A0A7M1AWW5 | |
Candidatus Sulfurimonas marisnigri | A0A7S7RQV3 | |
Sulfurimonas aquatica | A0A975AYP7 | |
Sulfurimonas autotrophica | E0UQ91 | |
Sulfurospirillum deleyianum | D1B011 | |
Nitratifractor salsuginis | E6X0A3 | |
Sulfurovum lithotrophicum | 285 | A0A7U4RQ57 |
When available, UniProt entries include a cross-reference link to KEGG, a database that provides genomic maps of numerous species, allowing one to trace back from a protein to the genomic location of its encoding gene. Of the 49 previously identified DciA proteins, 24 are linked to KEGG, corresponding to 22 bacterial species, see Supplemental file 3. These include Malaciobacter halophilus and Candidatus Campylobacter infans, which, as previously mentioned, harbor two dciA genes. Of these 24 dciA genes, 17 appear to be co-localized with uvrC (encoding the excinuclease ABC subunit C), see Table below. Interestingly, among these 17 dciA genes, two belong to the genus Helicobacter, namely Helicobacter bizzozeronii (strain CIII-1) and Helicobacter hepaticus (strain ATCC 51449 / 3B1).
In summary, co-localization of dciA and uvrC is a feature of numerous Campylobacterales.
In H. pylori, uvrC co-localizes with dciA
Among the 77 Campylobacterales species previously identified (see previous section), two correspond to H. pylori: H. pylori strain ATCC 700392 / 26695 and H. pylori strain G27. Both strains appeared to lack DciA (no DUF721 annotated protein). Given that dciA has been shown to co-localize with uvrC in numerous Campylobacterales (see previous section), I examined the genomic context of uvrC in both H. pylori strains.
In H. pylori strain ATCC 700392 / 26695 and H. pylori strain G27, uvrC is identified as UVRC_HELPY and UVRC_HELPG, respectively, and is encoded by the genes HP_0821 and HPG27_780, respectively. Using the KEGG link provided by UniProt to trace the protein back to its gene, uvrC appears to be located on the direct strand at positions 873393 to 875177 (https://www.genome.jp/dbget-bin/www_bget?hpy:HP_0821) and 844372 to 846156 (https://www.genome.jp/dbget-bin/www_bget?hpg:HPG27_780), respectively. Adjacent to uvrC on the same strand, I identified the genes HP_0820 (https://www.kegg.jp/entry/T00008:HP_0820) and HPG27_779 (https://www.kegg.jp/entry/T00777:HPG27_779) at positions 872931 to 873392 and 843910 to 844371, respectively, encoding proteins of 153 amino acids with the following primary sequences:
MEQNIFSLLIQKKSYKKLETLLKLKKLKVFMPLSLQENLLFIFIKDSKLLFAFKDIWASKEFNQRFAKEISHFLNTQGHAYGFDGLNGLEILGYVPKDALKKSNFYAPIKKQARFFRPSALGLFHNPIKDARLHECFEKARALIHYQRSFFEE
and
MEQNIFSLLIQKKSYKKLETLLKLKKLKVFMPLSLQENLLFIFIKDSKLLFAFKDLWASKEFNQRFAKEISHFLNTQGHAYGFDGLNGLEILGYVPKDALKKANFYAPIKKQARFFRPSALGLFHNPIKDARLHECFEKARALIHYQRSFFEE
respectively, see part A of Figure 1 for HP_0820/O25498_HELPY.
In UniProt, HP_0820 and HPG27_779 encode the proteins O25498_HELPY and B5Z7I5_HELPG, respectively, which are described as uncharacterized proteins with neither InterPro nor Pfam annotations. These two proteins share 99% sequence identity and differ by two amino acid mismatches
I began by reannotating these proteins using HHpred. Running HHpred (with default parameters) against Pfam-A_37 (the latest Pfam release, June 2024, containing 21979 families), I found that both O25498_HELPY and B5Z7I5_HELPG harbor the Pfam PF05258/DUF721 domain with a probability of 0.986, see Supplemental files 4 and 5, see part B of Figure 1 for HP_0820/O25498_HELPY. This domain spans residues 2 to 79. According to the HHpred documentation, a probability exceeding 0.95 indicates near-certain homology.
Subsequently, I examined the AlphaFold-predicted structures of O25498_HELPY (https://www.uniprot.org/uniprotkb/O25498/entry#structure) and B5Z7I5_HELPG (https://www.uniprot.org/uniprotkb/B5Z7I5/entry#structure), as provided (if available) within the UniProt protein entries. Both structures show very high or high confidence for most residues, with the N-terminal alpha-helix and the linker region exhibiting lower confidence. I then observed that these two AlphaFold structures contained the canonical structural DciA domain architecture, which consists of one alpha helix, followed by two beta sheets, a third alpha helix, and a final beta sheet (Chan-Yao-Chong et al., 2020).
I therefore concluded that HP_0820 and HPG27_779 are the dciA genes of H. pylori strains ATCC 700392 / 26695 and G27, respectively.
Discussion
Despite the apparent absence of dciA from the H. pylori genome (Blaine et al., 2023), the unique characteristics of its replicative helicase (HpDnaB), when compared to other bacterial replicative helicases (Stelter et al., 2012), prompt me to propose that dciA is still present but has co-evolved with HpDnaB, making it difficult to detect. To test this hypothesis, I decided to use ‘genomic synteny’, which assumes that during evolution, while genomes can change, some gene arrangements are preserved.
With the growing availability of proteome and genome data, I narrowed my focus to the 77 Campylobacterales species, the order to which H. pylori belongs, available as ‘reference proteomes’ in UniProt (UniProt Consortium, 2024). These 77 Campylobacterales proteomes include, in particular, those of two Helicobacter pylori strains: ATCC 700392 / 26695 and G27. Identifying DciA using the standard DciA Pfam domain signature method, I found that 46 of the 77 Campylobacterales proteomes host a DciA protein. To trace back from proteins to their encoding genes, I relied, when available, on cross-referencing links to KEGG (Kanehisa et al., 2024), a database hosting genomic maps of many species. Among the aforementioned 46 species, 22 are stored in KEGG, offering the opportunity to examine the genomic context of dciA. I then observed that, frequently, dciA is found to co-localize with the uvrC gene, i.e., the excinuclease ABC subunit C gene (see Results section ‘Genomic co-localization of the dciA and uvrC genes is a feature of numerous Campylobacterales’). Interestingly, this pattern is observed in particular in two close relatives of Helicobacter pylori, namely Helicobacter bizzozeronii (strain CIII-1) and Helicobacter hepaticus (strain ATCC 51449 / 3B1).
I then returned to H. pylori to examine the genomic context of its uvrC gene. As previously mentioned, two Helicobacter pylori strains are stored in UniProt as ‘reference proteomes’, namely Helicobacter pylori strain ATCC 700392 / 26695 and Helicobacter pylori strain G27. I focus here only on Helicobacter pylori strain ATCC 700392 / 26695, as similar results are obtained for the other strain (see Results section).
The ordered locus name of the uvrC gene in H. pylori strain ATCC 700392 / 26695 is HP_0821. Adjacent to HP_0821, I identified a gene, HP_0820, located on the same strand, encoding a 153-amino-acid-long protein stored as O25498_HELPY in UniProt, see part A of Figure 1. The UniProt ‘Family & Domains’ section provides no annotation for O25498_HELPY. This result indicates that the InterProScan tool (Blum et al., 2025) failed to detect any domain in O25498_HELPY when queried against its integrated member database signatures (e.g., Pfam, PROSITE, etc.). I then submitted HP_0820 to HHpred (Gabler et al., 2020), a profile-profile comparison tool for remote homology detection, which identified the DciA-characteristic Pfam DUF721 domain (probability > 0.95) in HP_0820, see part B of Figure 1. This was further supported by AlphaFold’s 3D prediction (Jumper et al., 2021) of HP_0820, which revealed the presence of the canonical DciA domain structure, see part C of Figure 1.
Taken together, my results strongly suggest that the proteins encoded by HP_0820 in H. pylori strain ATCC 700392 / 26695 and HPG27_780 in H. pylori strain G27 are homologous to DciA (see Results section ‘In H. pylori, uvrC co-localizes with dciA’). This finding significantly challenges the two current models of chromosome replication initiation in H. pylori, as explained below.
The first model suggests that H. pylori encodes a gene, HP_0897, which putatively functions as a replicative helicase operator/loader (Verma et al., 2016; Kumar et al., 2019). In UniProt, HP_0897 corresponds to a protein with ID O25557_HELPY. When submitted to HHpred, analysis revealed that HP_0897 harbors the Pfam DUF721 domain, albeit with a reduced probability (0.884) compared to HP_0820. A visual examination of the HP_0897 AlphaFold structure indicates that, similar to HP_0820, it exhibits the canonical DciA domain structure. The presence of this domain likely explains HP_0897’s interaction with the replicative helicase DnaB, mirroring the behavior of DciA in several well-studied bacteria (Caulobacter crescentus (Ozaki et al., 2022), Mycobacterium tuberculosis (Mann et al., 2017), Pseudomonas aeruginosa (Brézellec et al., 2016), and Vibrio cholerae (Marsin et al., 2021)). Interestingly, HP_0820 also interacts with DnaB in Helicobacter pylori (Häuser et al., 2014). However, a significant difference exists between HP_0820 and HP_0897: HP_0820, but not HP_0897, is an essential gene (Salama et al., 2004), a key characteristic of DciA (Blaine et al., 2022). Nevertheless, while focusing on the growth of knockouts in rich media is one way to identify essential genes, gene persistence is another approach to assess a gene’s essentiality (Fang et al., 2005). I thus investigated the gene persistence of both HP_0820 and HP_0897. KEGG currently stores 61 complete H. pylori genomes (including strains ATCC 700392 / 26695 and G27). Using the KEGG DBsearch function (which performs a BLASTP of a protein query against the database of all genes stored in KEGG), HP_0820 was found to be persistent—meaning it is present in all 61 H. pylori strains examined (see Supplemental file 6)—whereas HP_0897 is not. Specifically, HP_0897 is found in only 45 out of the 61 strains (see Supplemental file 7). In conclusion, HP_0897 is neither essential in H. pylori strain ATCC 700392 / 26695 nor persistent in the set of 61 H. pylori strains examined, unlike HP_0820. Taken together, these results strongly suggestthat DciA/HP_0820, rather than HP_0897, assists H. pylori DnaB at the onset of chromosome replication initiation.
The second model, originally proposed prior to the discovery of HP_0897 (Blaine et al., 2023), posits that H. pylori DnaB can self-load onto DNA without requiring a ‘dedicated’ helicase loader, a hypothesis supported by a study demonstrating HpDnaB’s ability to bypass DnaC in E. coli and function as an active replicative helicase (Soni et al., 2005). However, experiments carried out in the DciA-containing bacterium Vibrio cholerae El Tor strain E7946 showed that V. cholerae DnaB can load itself onto DNA in vitro, and that V. cholerae DciA stimulates this function, resulting in increased DNA unwinding (Marsin et al., 2021). Therefore, the ability to self-load onto DNA might be a common feature of replicative helicases in DciA-containing organisms.
Overall, my work convincingly suggests that dciA is present in the Helicobacter pylori genome and likely functions as a replicative helicase loader, significantly challenging current models of chromosome replication initiation for this organism. Furthermore, it highlights the limitations of the current methodology used to identify DciA. Moreover, as pointed out by Radford et al. (2023), in the development of novel antibiotics, DNA replication remains a largely underexplored area, even though replisomal proteins are clearly attractive therapeutic targets. This potential can only be realized if the proteins involved in chromosome replication are well-characterized. My work suggests that this criterion is now likely met for Helicobacter pylori.
Numerous questions remain to be addressed to fully elucidate replication initiation in Campylobacterales. While fruitful in the case of H. pylori, my hypothesis that DciA has co-evolved with HpDnaB, leading to the loss of its classical Pfam domain signature, requires further verification across the entire Campylobacterales order. Alternatively, as previously shown, dciA might have been repeatedly replaced by phage helicase loaders. Both possibilities warrant further investigation.
Acknowledgements
This work is dedicated to the memory of Jean-Luc Férat, who has passed away. I met Jean-Luc in 2000 and began working with him a few months later. Our work on the Dam methyltransferase led to the development of methodologies that paved the way for the discovery of dciA several years later. Throughout this period, Jean-Luc’s contributions were crucial. I will deeply miss his insightful ideas, intuition, and discussions.
I am grateful to the recommender, Rafael Laso-Pérez, and the two referees for their comments and suggestions, which significantly improved the quality of the final submission. Preprint version 3 of this article has been peer-reviewed and recommended by Peer Community In PCI Genomics (https://doi.org/10.24072/pci.genomics.100430; Laso-Pérez, 2026).
Funding
The author declares that he has received no specific funding for this study.
Conflict of interest disclosure
The author declares that he complies with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data, scripts, code and supplementary information availability
Supplementary information is available online (https://doi.org/10.5281/zenodo.17592664; Brézellec, 2025).