
 CC-BY 4.0
CC-BY 4.0
Introduction
The genus Trichoderma (Ascomycota, Hypocreaceae) includes fungi widely recognized for their biocontrol potential in agriculture. To date, 225 Trichoderma genomes are publicly available on NCBI, but only a few have been assembled to completion: the reference genomes of Trichoderma atroviride, Trichoderma asperellum, Trichoderma asperelloides, Trichoderma simmonsii, and Trichoderma semiorbis. Complete genomes are also available for Trichoderma reesei and Trichoderma virens, although these are not designated reference genomes. High-quality assemblies are particularly valuable for comparative genomics and for identifying genes involved in pathogen suppression, secondary metabolism, and other biocontrol-related functions (Li et al., 2021; Schalamun & Schmoll, 2022; Rosolen et al., 2023).
Trichoderma gamsii is a ubiquitous fungus found in diverse environments, where it can exhibit psychrotolerance and broad pH tolerance (Migheli et al., 2009; Rinu et al., 2014). This species is particularly interesting due to its ability to suppress pathogens through enzymatic activity and the release of volatile organic compounds (Anees et al., 2010; Chen et al., 2016; Galletti et al., 2020; Bedine Boat et al., 2020; Valan Arasu et al., 2023; Marik et al., 2025). Additionally, it promotes plant health by inducing systemic resistance and stimulating plant growth (Anees et al., 2010; Rinu et al., 2014; Chen et al., 2016; Galletti et al., 2020; Valan Arasu et al., 2023). These characteristics highlight the potential of T. gamsii for biotechnological applications, underscoring the importance of comprehensive genomic data for further investigation of its biocontrol abilities. To date, the best public genomic sequence available for this species belongs to the strain T6085, which is known for its ability to control Fusarium head blight on wheat (Baroncelli et al., 2016).
In this study, we performed de novo sequencing of T. gamsii T035 genome. This strain exhibits strong antagonistic activity against multiple plant pathogens, making it a promising model to explore T. gamsii biocontrol potential. Here, we present a high-quality, near chromosome-level genome assembly and its functional annotation. With only 16 sequences, seven of which corresponding to near chromosome-scale nuclear scaffolds, this assembly represents the most contiguous T. gamsii genome available to date and provides a valuable resource to support future comparative and applied studies.
Material and methods
Sample information
Trichoderma gamsii strain T035 was isolated in 2019 from carrot-cultivated soil in Brain-sur-l’Authion, France (GPS coordinates: latitude 47.473936; longitude −0.390684) (Figure 1). Sampling and isolation were performed following the method described by Chateau et al. (2024). Briefly, 5 g of rhizosphere soil were suspended in 50 mL of sterile water. Serial dilution–extinction was carried out on Potato Dextrose Agar (PDA; Grosseron, 39 g L⁻¹) supplemented with streptomycin (500 mg L⁻¹). Cultures were incubated in darkness at 20 °C for seven days. Strain T035 was subsequently transferred to Malt Agar medium (MA; Grosseron, 20 g L⁻¹ bacteriological malt extract, 15 g L⁻¹ bacteriological agar type A) and maintained under the same growth conditions. For long-term preservation, mycelial explants were stored at −80 °C in 30% glycerol solution as a cryoprotectant.
Trichoderma atroviride strain I1237 was obtained from the commercial product TRI-SOIL® (Agrauxine, Beaucouzé, France).
Pathogenic strains were sourced either from our laboratory fungal collection (IRHS, FungiSem Team, Beaucouzé, France) or from the CIRM-CFBP collection (Beaucouzé, France). The twelve filamentous pathogens belong to Rhizoctonia solani (PAT-002/PAT-006/PAT-009), Globisporangium sp. (PAT-013/PAT-018/PAT-019), Stagonosporopsis valerianellae (PAT-023/PAT-024/PAT-025), and grapevine trunk disease fungi, namely Phaeomoniella chlamydospora (PAT-026), Diplodia seriata (PAT-027), and Neofusicoccum parvum (PAT-028). The two bacterial strains used in this study were Acidovorax valerianellae (PAT-034) and Xanthomonas campestris (PAT-035).They are further described in the Supplementary Table S1.

Figure 1 - Colony morphology of Trichoderma gamsii T035 on PDA after 7 days of incubation, with bottom (left) and top (right) views of the plate (A). Confrontation assay between T. gamsii T035 (bottom-right) and Rhizoctonia solani PAT-009 (top-left) (B). Hyphal coiling of T. gamsii T035 (Tg) around R. solani PAT-009 (Rs) (scale bar = 50 µm) (C).
In vitro control of filamentous pathogens
The two Trichoderma isolates were evaluated for their mycoparasitic activity against the twelve pathogenic fungal and fungal-like isolates previously described using the dual culture assay (Bunbury-Blanchette & Walker, 2019). Square Petri dishes (120 × 120 mm) containing PDA medium were inoculated with two 5 mm mycelial plugs: one obtained from a pathogen colony and the other from a Trichoderma colony previously grown on PDA under the conditions described above. The plugs were placed at opposite corners of the plate, with the pathogen inoculated two days before the antagonist. Each condition was replicated three times, and the entire experiment was repeated independently three times.
Plates were incubated in darkness at 20 °C for 20 days. Mycoparasitic ability was visually assessed using a scale adapted from Pascouau et al. (2023): 0 = <5% of pathogen colony overgrown by Trichoderma; 1 = 5–25% overgrown; 2 = 25–50% overgrown; 3 = 50–75% overgrown; 4 = >75% overgrown (Figure S1).
Statistical analyses were performed in R v4.3.1. Data were analyzed using a non-parametric analysis of variance (Wilcoxon signed-rank test), and p-values were adjusted for multiple testing using the false discovery rate (FDR) method.
In vitro control of bacterial pathogens
Two Trichoderma isolates were assessed for their antibiosis capacity against the bacterial isolate PAT-034 (A. valerianellae) and PAT-035 (X. campestris) following the protocols of Dennis & Webster (1971) and Lee et al. (2012), with modifications. For each Trichoderma isolate, 250 mL Erlenmeyer flasks containing 50 mL of Potato Dextrose Broth (PDB, Grosseron, 24 g L⁻¹, pH 5.2) were inoculated with five mycelial plugs (5 mm diameter) taken from 7-day-old PDA cultures. Cultures were incubated at 20 °C on a rotary shaker (100 rpm) for 7 days and subsequently filtered through sterile 0.2 μm syringe filters (ClearLine®, Dutscher) to obtain culture filtrates.
The effect of the filtrates on bacterial growth was evaluated using a spectrophotometric microplate assay (96-well plates, SPECTROstar, BMG Labtech). Autoclaved Lysogeny Broth (LB, Grosseron, 24 g L⁻¹) was inoculated with 1% (v/v) of a bacterial suspension adjusted to OD = 0.05, supplemented with 10% (v/v) of Trichoderma culture filtrates, in a final volume of 300 μL per well. Filtrates obtained from non-inoculated PDA disks were incubated under the same conditions and served as controls. Plates were incubated at 25 °C for 33 h with intermittent shaking (5 min at 200 rpm every 10 min), and absorbance at 600 nm was recorded every 10 min.
Each condition was tested in triplicate, and three independent experiments were conducted. The area under the growth curve was calculated as described by Joubert et al. (2010) and Warringer & Blomberg (2003). Data were analyzed using analysis of variance (ANOVA) followed by Tukey’s multiple comparisons test (α = 0.05).
Genome-wide relatedness among Trichoderma species
A global comparison of Trichoderma genomes was performed using MashTree v1.2 (Katz et al., 2019). Genome assemblies of T. gamsii and representative assemblies from other Trichoderma species were retrieved from NCBI RefSeq (accessions listed in Supplementary Table S2).
Pairwise genomic distances were computed with Mash (genome size parameter set to 40 Mbp, sketch size = 20,000). The distance matrix was used to infer a neighbor-joining tree with MashTree. The obtained tree was exported in Newick format, and later visualized and annotated using MEGA12 (Kumar et al., 2024) (Figure 2).
DNA extraction
High-molecular-weight genomic DNA was extracted from a 7-day-old culture of T. gamsii T035 grown on a pectocellulosic membrane placed on PDA medium. Tissues were frozen with liquid nitrogen and bead-milled. DNA extraction followed an adapted Möller protocol: RNase treatment was performed immediately after proteinase K digestion, and DNA was resuspended in 10 mM Tris (pH 8) instead of TE buffer, as EDTA can interfere with Nanopore sequencing (Möller et al., 1992). The A260/280 and A260/230 ratios were 1.85 and 2.3, respectively. DNA concentration was measured using the quBit to 27 ng/µL, and its integrity was confirmed by electrophoresis gel, where the majority of the gDNA was above 10 kilobases (kbp) in length.
Oxford Nanopore Technologies (ONT) sequencing
The sequencing library was prepared using the Native Barcoding Kit 96 V14 (SQK-NBD114.96) with 1.5 µg of high-molecular-weight genomic DNA as described in the kit protocol. We performed de novo sequencing of T. gamsii T035 genome using Oxford Nanopore Technologies’ MinION device M1kD on the flowcell FLO-MIN114 with R10.4 pores (Lu et al., 2016).
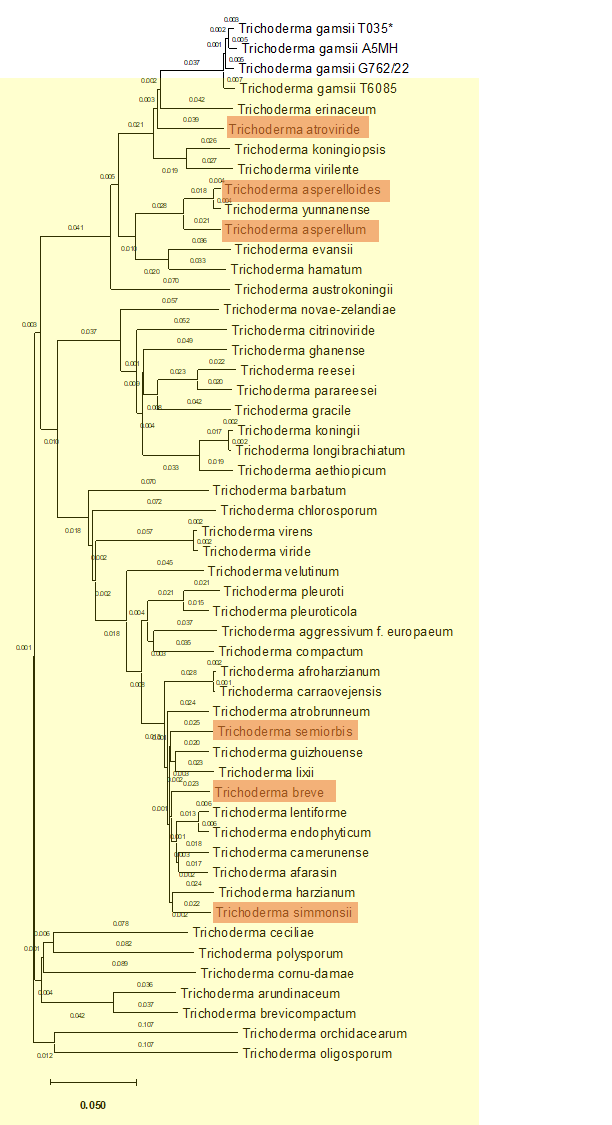
Figure 2 - Genetic distance-based tree of Trichoderma genomes.
The analysis included all reference Trichoderma genomes available in NCBI (yellow) as well as all known T. gamsii genomes. The genome obtained in this study is indicated with an asterisk. Complete genomes are highlighted in orange.
Genome assembly
Reads were basecalled using the super accuracy (SUP) mode. The long-read assemblies were performed using both Canu v2.2 (Koren et al., 2017) and Flye v2.9.6 (Kolmogorov et al., 2019). The less fragmented assembly was obtained using Flye, with reads filtered before assembly where reads shorter than 1,000 bp or with a quality score below Q10 were removed (De Coster et al., 2018). The assembly underwent three additional polishing steps using Racon v1.5.0 (Vaser et al., 2017). Contigs were scaffolded with RagTag v2.1.0 (Alonge et al., 2022) using T. atroviride GCF_020647795.1 as reference. Assembly quality was later assessed with BUSCO 5.4.4 (fungi_odb10 database) and compared to the reference T. gamsii genome (RefSeq: GCF_001481775.2) using QUAST v5.3.0 (Mikheenko et al., 2023; Tegenfeldt et al., 2025) (Figure 3 and Table 1). Additionally, k-mer spectra plots were generated from Canu-corrected reads (Koren et al., 2017) using Jellyfish v2.2.10 (Marçais & Kingsford, 2011), and Genoscope 2.0 (Ranallo-Benavidez et al., 2020) (Figures S2 - S5). Genome representation was performed using BlobToolKit v4.0.7 (Challis et al., 2020) (Figure 3).
Table 1 - Comparison of global genome metrics of Trichoderma gamsii genomes. The asterisk indicates the actual reference genome of T. gamsii in NCBI. The “m” indicates the mitochondrial sequences.
|
Assembly statistics |
Trichoderma gamsii strain t6085* |
Trichoderma gamsii strain a5mh |
Trichoderma gamsii strain g762/22 |
Trichoderma gamsii strain t035 (this study) |
|---|---|---|---|---|
|
Submitted genbank assembly |
GCA_001481775.2 |
GCA_002894205.1 |
GCA_049996675.1 |
|
|
Genome size |
37.9 Mbp |
38.5 Mbp |
38 Mbp |
38.8 Mbp |
|
Total ungapped length |
37.9 Mbp |
38.5 Mbp |
38 Mbp |
38.8 Mbp |
|
Number of scaffolds |
172 |
292 |
499 |
8 (7+1m) |
|
Scaffold n50 |
697.4 kbp |
594.5 kbp |
285.3 kbp |
7.2 Mbp |
|
Scaffold l50 |
18 |
20 |
44 |
3 |
|
Number of contigs |
216 |
517 |
499 |
30 |
|
Contig n50 |
534 kbp |
299.2 kbp |
285.3 kbp |
3.096 Mbp |
|
Contig l50 |
23 |
42 |
44 |
5 |
|
Gc percent |
49 |
48.5 |
48.5 |
48.4 |
|
Genome coverage |
50x |
90x |
35x |
25.4x |
|
Assembly level |
Scaffold |
Scaffold |
Contig |
Scaffold |
Annotation
Structural annotation was performed using the deep-learning tool Helixer (Holst et al., 2025). Telomeres were detected with TIDK v0.2.65, allowing the identification of 1,097 repeats of the canonical motif AAAAAAT (Hofstatter et al., 2022). To confirm telomere locations and identify centromeres, a sliding window analysis was performed using bedtools v2.31.1 and RepeatMasker v4.2.2 to detect regions with high repeat density, low GC content, and low gene density. Centromeric and telomeric regions were then manually curated by integrating overlapping signals from these analyses. The distribution of transposable elements (TEs) across the genome was assessed using EDTA v2.2.2 (Su et al., 2021) (Supplementary Table S3). RIP regions were determined in TEs using The RIPper (http://theripper.hawk.rocks) (van Wyk et al., 2019).
Functional annotation was carried out with EggNOG-mapper v2 (eggNOG database version 5.0.2) (Cantalapiedra et al., 2021) (Supplementary Table S4), signal peptides were predicted using DeepSig (Version 1.2.5) (Savojardo et al., 2018) (Supplementary Tables S5-6), secondary metabolite (SM) clusters were identified using antiSMASH 8.0 (fungal mode) (Blin et al., 2025) (Supplementary Table S7), and carbohydrate-active enzymes (CAZymes) were identified using dbCAN3 (dbCAN HMMdb v12) (Zheng et al., 2023) (Supplementary Table S8). Only high-confidence enzymes were kept in this study, defined as matches detected by all tested tools: HMMER: dbCAN (E-value < 1e-15, coverage > 0.35), DIAMOND: CAZy (E-value < 1e-102), and HMMER: dbCAN-sub (E-value < 1e-15, coverage > 0.35). All annotation results are synthesized in the Supplementary Table S9. A comparison of secondary metabolite clusters, proteins with signal peptide, and carbohydrate-active enzymes was performed on T. gamsii genomes as well as the complete reference genomes of Trichoderma available on NCBI (Supplementary Table S2).

Figure 3 - Snailplot generated with BlobToolKit showing assembly statistics for the Trichoderma gamsii T035 genome. The assembly has a total length of 38,792,561 bp with a maximum scaffold length of 9,818,134 bp (in red). N50 and N90 sequence lengths are 7,178,191 bp and 4,032,368 bp, indicated by orange and pale-orange arcs, respectively. The grey spiral represents scaffold lengths in descending order on a log scale. BUSCO analysis indicates 98.8% completeness (top right). The outer ring shows base composition (GC: 48.4%, AT: 51.6%, N: 0.0%). It’s important to note that BlobToolKit reports 0.0% N, as the gaps introduced by RagTag during the scaffolding are synthetic and not associated with a raw sequence.
Trichoderma genome comparison
To identify syntenic regions between T. gamsii and Trichoderma atroviride (Figure 6B), as well as among the different T. gamsii genomes (Figure S6), comparative genomic and synteny analyses were performed using the JCVI toolkit v1.4.16 (Tang et al., 2024). Orthologous genes were identified based on protein sequence similarity using BLAST, and conserved syntenic regions were inferred from the genomic distribution and colinearity of these orthologs along chromosomes. Syntenic blocks were filtered to retain only robust regions supported by at least 30 consecutive colinear orthologous genes. Filtered syntenic relationships were visualized using JCVI-based karyotype representations.
Results
Biocontrol assays
The biocontrol potential of T. gamsii strain T035 was assessed through its parasitism and antibiosis capacities. Strain T035 displayed greater parasitic activity than the commercial strain I1237 against eight of the twelve tested filamentous plant pathogens (Figure 4A). Notably, while I1237 exhibited antibiosis activity against both Acidovorax valerianellae and Xanthomonas campestris, strain T035 was more than twice as effective (Figure 4B).
An intriguing pattern emerged in the parasitism assays with Globisporangium species: strain T035 was particularly effective against isolate PAT-018, but showed markedly reduced activity against the two other Globisporangium strains tested (Figure 4A).

Figure 4 - Evaluation of biocontrol capacities of T. gamsii strain T035 compared with a commercial Trichoderma strain (I1237). (A) Radar plot showing the parasitic capacity of both strains against twelve isolates from four different filamentous plant pathogens. Scores range from 0 (no colonization) to 4 (complete colonization). Asterisks indicate significant differences between strains (Wilcoxon signed-rank test; p-value 0.05). (B) Boxplot showing the growth of two phytopathogenic bacteria under control conditions (grey) or after exposure to culture filtrates of strain I1237 (pink) and T. gamsii T035 (blue). Different letters indicate significant differences according to Tukey’s multiple comparisons test (α = 0.05).
Genome sequencing and assembly results
ONT sequencing of T. gamsii strain T035 generated 196,604 reads with a N50 read length of 6,669 bp, yielding approximately 965 Mbp of sequence data, which corresponds to around 25X genome coverage (Figure 5). The median read quality and length were respectively 21.8 and 3,217 bp.
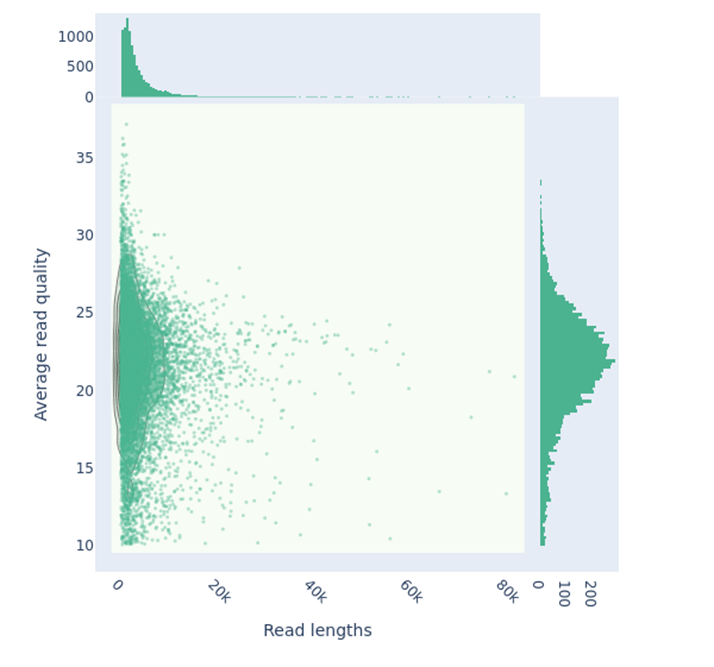
Figure 5 - Scatterplot of Nanopore reads for T. gamsii strain T035, showing read length versus quality score, generated with NanoPlot (kde plot). A total of 196,604 reads were analyzed, representing approximately 965 Mbp of sequence data. The read length N50 is 6,669 bp, with a median read length of 3,217 bp, while the median read quality is 21.8.
The initial Flye assembly of T. gamsii T035 produced 30 contigs (N50 = 3.1 Mbp and N90 = 1.3 Mbp), which were scaffolded into 16 sequences (N50 = 7.18 Mbp and N90 = 4.03 Mbp ; 7 nuclear chromosomes, 5 unassigned contigs and 4 mitochondrial sequences), which introduced 3.09 N’s per 100 kbp (Figure 3, Figure 6 and Table 1). The total assembly length is of 38.8 Mbp, with a GC content of 48.44% and a largest contig of 7.09 Mbp. Assembly completeness, assessed with BUSCO, indicated 98.8% complete universal genes (complete and single-copy: 98.3%; complete and duplicated: 0.5%; fragmented: 0.4%; missing: 0.8%) against the fungi_odb10 database (Figure 3) (Tegenfeldt et al., 2025). Key genome metrics for T035 are compared with other published T. gamsii genomes in Table 1, and synteny analysis between T035, T6085, and A5MH highlights the high fragmentation of T6085 and A5MH relative to T035 (Supplementary Figure S2).
Genomic structure
The T. gamsii T035 genome is composed of seven large sequences corresponding to chromosomes. These chromosomes are well defined, as most of them are flanked by telomeric regions, except for chromosome CP084936.1 for which one telomeric region was missing (Figure 6A). Notably, chromosome CP084937.1 is almost twice as long in T. gamsii T035 as in T. atroviride P1. This size difference is likely attributable to an interchromosomal translocation from chromosome CP084938.1 to chromosome CP084937.1 (Figure 6B). As a consequence, chromosome CP084938.1 is markedly shorter in T. gamsii T035 than in T. atroviride P1 (2.22 Mbp versus 5.27 Mbp). This reduction in size may explain why no centromeric region was identified on chromosome CP084938.1 in T. gamsii T035 (Figure 6A).

Figure 6 - Complete sequences of the seven chromosomes of T. gamsii obtained in this study, compared with the reference genome of T. atroviride P1. (A) Chromosomal structures of T. gamsii. Telomeres are shown in blue and centromeres in red. One telomere was not detected on chromosome CP084936.1, and the centromere was not detected on chromosome CP084938.1. Chromosome sizes for T. gamsii T035 and the T. atroviride reference genome P1 are shown on the right. (B) Synteny analysis between the genomes of T. gamsii T035 and T. atroviride P1 shows that the expansion of chromosome CP084937 in T. gamsii T035 is due to an interchromosomal translocation from chromosome CP084938.
Genome structural annotation and annotation quality control
Structural annotation was performed using the deep-learning tool Helixer (Holst et al., 2025), which predicted 13,036 protein-coding genes. Compared to the previously published reference genome of T. gamsii strain T6085, annotated using MAKER2 (Holt & Yandell, 2011), the annotation of strain T035 identified more than 2,000 additional protein-coding genes (Table 2).
Recent studies have reported that deep-learning-based predictors may occasionally generate unsupported gene models (Lapalu et al., 2025). Therefore, part of the increase in predicted gene number may reflect differences between annotation pipelines rather than true biological differences between strains.
TE analysis revealed that 3.39% of the T035 genome is composed of transposable elements (Table 3 and Supplementary Table S3). The most abundant superfamilies were LTR retrotransposons of the Gypsy type (0.52%), helitrons (0.58%), and TIR elements of the CACTA and Mutator families (0.17% each). Other TE classes, including LINEs and SINEs, were only marginally represented (<0.1%).
Table 2 - Genome annotation metrics comparing T. gamsii (strain T035) from this study with the reference genome T. gamsii (strain T6085) from Baroncelli et al. (2016) as well as with the completed genomes available on NCBI. Asterisks indicate values reported in Baroncelli et al. (2016); values without asterisks were generated in this study. Signal peptides were detected using DeepSig, SM clusters were predicted using antiSMASH, and CAZymes were identified using dbCAN3.
|
Trichoderma gamsii T6085 |
Trichoderma gamsii T035 |
Trichoderma atroviride P1 |
Trichoderma asperellum FT101 |
Trichoderma asperelloides X01119 |
Trichoderma simmonsii GH-Sj1 |
Trichoderma semiorbis FJ059 |
Trichoderma breve T069 |
|
|---|---|---|---|---|---|---|---|---|
|
Rreference genome (Baroncelli et al., 2016) |
Genome from this study |
NCBI RefSeq |
NCBI RefSeq |
NCBI RefSeq |
NCBI RefSeq |
NCBI RefSeq |
NCBI RefSeq |
|
|
Predicted protein-coding genes |
10,944* |
13,036 |
13327 |
12041 |
11096 |
13296 |
10247 |
11600 |
|
Signal peptide |
933 (1356*) |
1,046 |
892 |
908 |
891 |
1046 |
896 |
1006 |
|
Isocyanide |
2 |
2 |
1 |
2 |
1 |
0 |
0 |
0 |
|
fungal-RiPP |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
0 |
|
NRPS |
14 |
11 |
13 |
13 |
11 |
17 |
14 |
16 |
|
PKS |
11 |
11 |
11 |
10 |
8 |
17 |
17 |
19 |
|
Terpene |
10 |
12 |
13 |
12 |
12 |
15 |
13 |
15 |
|
Isocyanide-nrp |
3 |
3 |
2 |
2 |
2 |
1 |
1 |
1 |
|
Isocyanide/NRPS |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
1 |
|
NRPS/indole |
0 |
0 |
0 |
0 |
0 |
2 |
0 |
1 |
|
NRPS/Terpene |
0 |
0 |
0 |
1 |
1 |
0 |
1 |
0 |
|
PKS/NRPS |
4 |
4 |
4 |
6 |
6 |
9 |
8 |
8 |
|
PKS/Terpene |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
|
Isocyanide-nrp/Isocyanide/Terpene |
0 |
0 |
1 |
1 |
1 |
0 |
0 |
0 |
|
NRPS/PKS/terpene |
1 |
1 |
1 |
1 |
1 |
1 |
0 |
0 |
|
Total of detected SM clusters |
45 |
44 |
46 |
48 |
44 |
65 |
56 |
62 |
|
Glycoside hydrolase (GH) |
250 |
257 |
220 |
251 |
242 |
236 |
213 |
224 |
|
Glycosyltransferase (GT) |
87 |
88 |
75 |
86 |
81 |
77 |
92 |
69 |
|
Polysaccharide Lyase (PL) |
10 |
10 |
9 |
10 |
9 |
7 |
4 |
8 |
|
Carbohydrate Esterase (CE) |
19 |
20 |
17 |
19 |
19 |
19 |
14 |
19 |
|
Carbohydrate-Binding Module (CBM) |
33 |
33 |
10 |
16 |
15 |
13 |
8 |
11 |
|
Auxiliary Activity (AA) |
45 |
44 |
36 |
39 |
43 |
43 |
38 |
43 |
|
Total of detected Cazyme |
444 |
452 |
367 |
421 |
409 |
395 |
369 |
374 |
Table 3 - Composition of transposable elements in Trichoderma gamsii strain T035 and in Trichoderma atroviride as determined with EDTA, RIP sequences were identified using The RIPper.
|
Class |
T. gamsii T035 |
T. atroviride P1 |
|||||
|---|---|---|---|---|---|---|---|
|
Nb of elements |
Nb of RIP |
% of sequence |
Nb of elements |
Nb of RIP |
% of sequence |
||
|
LINE |
Tad1 |
51 |
0 |
0.06% |
0 |
0 |
0.00% |
|
LTR |
Gypsy |
181 |
26 |
0.52% |
172 |
49 |
1.06% |
|
Copia |
0 |
0 |
0.00% |
1 |
0 |
0.00% |
|
|
Unknown |
78 |
15 |
0.26% |
9 |
0 |
0.00% |
|
|
SINE |
tRNA |
17 |
0 |
0.00% |
19 |
0 |
0.01% |
|
unknown |
15 |
1 |
0.00% |
0 |
0 |
0.00% |
|
|
TIR |
CACTA |
28 |
1 |
0.17% |
25 |
0 |
0.17% |
|
Mutator |
33 |
0 |
0.17% |
34 |
2 |
0.19% |
|
|
PIF_Harbinger |
8 |
0 |
0.03% |
2 |
1 |
0.01% |
|
|
Tc1_Mariner |
4 |
0 |
0.03% |
2 |
0 |
0.02% |
|
|
hAT |
1 |
0 |
0.00% |
3 |
0 |
0.01% |
|
|
nonTIR |
helitron |
23 |
1 |
0.58% |
9 |
0 |
0.24% |
|
rDNA |
45S |
13 |
0 |
0.01% |
0 |
0 |
0.00% |
|
Repeat_fragment |
- |
813 |
46 |
1.56% |
297 |
8 |
0.15% |
|
Total |
1265 |
90 |
3.39% |
573 |
60 |
1.87% |
|
Genome functional annotation
Functional annotation using EggNOG-mapper (Cantalapiedra et al., 2021) assigned functional annotations to 9,344 out of 13,036 predicted genes. The mating-type locus of T. gamsii T035 was identified using BLAST searches against known MAT proteins. Strong homology was detected with Trichoderma spinulosum MAT1-1-1, MAT1-1-2, and MAT1-1-3 protein sequences (best BLAST hits: 429 bits, e-value 1e-121; 549 bits, e-value 1e-157; and 271 bits, e-value 8e-074, respectively). No MAT1-2 genes were detected, indicating that T. gamsii T035 carries the MAT1-1 mating type.
Secretome prediction identified 1,046 proteins with signal peptides using DeepSig (Savojardo et al., 2018) (Table 2). For comparison, 933 signal peptide-containing proteins were identified using the same tool in the strain T6085, while 1,356 were previously reported using SignalP (Petersen et al., 2011; Baroncelli et al., 2016). Among the seven Trichoderma genomes analyzed in this study, T. gamsii T035 exhibited one of the highest number of predicted secreted proteins (Table 2).
SM biosynthetic gene clusters were predicted in both T. gamsii strains T035 and T6085, with 44 and 45 clusters identified, respectively (Table 2). The clusters comprised a globally similar distribution of types, including NRPS, PKS, terpene, isocyanide, and mixed NRPS/PKS clusters, reflecting comparable secondary metabolic potential in both strains. Notably, T. gamsii harbored fewer SM clusters than some other Trichoderma species, T. simmonsii in particular exhibited as high as 65 predicted SM clusters (Table 2).
Finally, CAZymes were identified with high confidence based on concordant predictions from HMMER, DIAMOND, and dbCAN_sub. A total of 452 and 444 CAZyme activities were detected in T. gamsii strains T035 and T6085, respectively (Table 2). Interestingly, compared with the other analyzed Trichoderma species, T. gamsii exhibited the highest number of predicted carbohydrate-active enzymes, whereas the other strains ranged from 367 to 421 CAZyme activities. This difference mainly resulted from an overrepresentation of carbohydrate-binding module (CBM) domains, which were two- to three-fold more abundant in T. gamsii than in the other tested Trichoderma species.
Discussion
In this study, we generated the most contiguous genome assembly of the biocontrol fungus T. gamsii to date, with scaffold N50 and L50 values of 7.18 Mbp and 3, respectively, compared to the previously best available assembly, which had an N50 of 697.4 kbp and an L50 of 18 (Table 2, Figure 3). The high quality of the assembly and annotation was further supported by a BUSCO analysis (Figure 3).
Biocontrol assays indicate that T. gamsii T035 exhibits higher antagonistic potential than a commercial Trichoderma strain, with significant inhibitory activity against several fungal and bacterial pathogens (Figure 4). The activity is particularly strong against the Basidiomycete Rhizoctonia solani and the Ascomycete Stagonosporopsis spp., whereas inhibition of wood-decay fungi is minimal. Activity against Globisporangium was isolate-dependent, suggesting a degree of host specificity (Figure 4A).
This specificity could involve host-selective toxins, carbohydrate-active enzymes, and/or secreted effectors, as reported for plant-pathogenic fungi (Hatta et al., 2002; Lyu et al., 2015; Ayukawa et al., 2021; Courtial et al., 2022). In Trichoderma, host-dependent regulatory pathways have also been described (Sharma et al., 2017), supporting the idea that T035’s biocontrol mechanisms may involve specialized attack pathways or differential interactions with host defenses. We therefore speculate that the observed specificity of T035 against Globisporangium spp. could involve specialized attack pathways, possibly mediated by host-specific toxins and/or effectors (Ramírez-Valdespino et al., 2019), or alternatively reflect differences in host defense mechanisms.
Interestingly, compared to other Trichoderma genomes, T. gamsii genomes seemed depleted in SM cluster but enriched in secreted proteins and CAZymes, particularly CBM-containing enzymes, which may indicate distinct antagonistic strategies among Trichoderma species. CBM-containing enzymes bind to cellulose and chitin and have been reported to be frequently located adjacent to chitinase genes in fungal genomes (Kubicek et al., 2011). These CBM domains are linked to the catalytic core of enzymes and enhance their efficiency, especially when acting on recalcitrant substrates or substrates present at low concentrations (Kubicek & Kubicek, 2016). Functional validation of the identified secondary metabolite clusters, CAZymes, and predicted secretome will be required to test these hypotheses.
Additionally, analysis of the global genome structure of T. gamsii T035 suggests an interchromosomal translocation from chromosome CP084938.1 to CP084937.1 when compared with T. atroviride P1 (Figure 6B). Confirmation of this rearrangement in other complete T. gamsii genomes will help determine when this event could have occurred.
Furthermore, the TE content of T035 is relatively low compared to other filamentous fungi, where TE content can go from 0,03% up to more than 80% (e.g., Fusarium graminearum and Blumeria graminis, respectively) (Ma et al., 2010; Kusch et al., 2024). However, Trichoderma species are generally TE-poor, with major TE classes typically representing 0.48 to 0.57% of the genome (Kubicek et al., 2011). In our study, TE content in T. atroviride was around 1.9%, while previous estimate for other T. atroviride genome was 0.49% (Kubicek et al., 2011). By comparison, T. gamsii T035 has a higher TE content (3.39%), most of which consists of highly degenerated repeat fragments. Moreover, the proportion of RIP-affected sequences is also higher in T. gamsii T035 (2.66%) than in T. atroviride P1 (1.77%).
Further studies across multiple Trichoderma species and strains will be required to better understand TE dynamics in this genus, as TE content is known to be highly strain-specific, as demonstrated in Leptosphaeria maculans (2.7–32.5%) (Grandaubert et al., 2014).
Overall, this study provides a high-quality genome assembly of T. gamsii T035, including detailed annotation of protein-coding genes, secondary metabolite clusters, carbohydrate-active enzymes, and the mating-type locus (MAT1-1). The T035 genome encodes 13,036 protein-coding genes, compared to 10,944 in the published reference genome of strain T6085. The genome has been deposited in ENA under BioProject PRJEB107169. This genomic resource, combined with functional assays, will support future studies aimed at elucidating the molecular basis of T. gamsii biocontrol activity and its potential applications in sustainable agriculture.
Acknowledgements
We thank the ANAN platform and the entire Nanopore Team, especially Muriel Bahut, Jessica Dittmer, Maryline Cournol, and Sandrine Balzergue, for the material and technical support during the genome sequencing. We acknowledge the GenOuest bioinformatics core facility (https://www.genouest.org) for providing the computing infrastructure.
Preprint version 4 of this article has been peer-reviewed and recommended by Peer Community In Genomics (https://doi.org/10.24072/pci.genomics.100458; Malagnac, 2026).
Funding
The authors declare that they have received no specific funding for this study.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data, scripts, code, and supplementary information availability
Data, supplementary figures and tables, scripts, and code are available online on GitHub and stored with Zenodo (https://doi.org/10.5281/zenodo.18417586; JustiCol, 2026).
T035 genomic data are available at http://www.ebi.ac.uk/ena/browser/view/PRJEB107169.