
 CC-BY 4.0
CC-BY 4.0
Introduction
In most poultry species, selection is carried out using individual cages in order to easily trace the pedigree of hatched chicks. Equipped with sloped floor allowing eggs to roll to the front of the cage where they are out of the hen’s reach and can be collected by the farmer, these cages gained popularity since their introduction in the early twentieth century (Arndt, 1931). Compared with a system where hens lay in a pen equipped with trap-nesting devices, broodiness and floor eggs are eliminated and eggs are cleaner. In addition, more birds can be housed in a given floor space.
Yet, in 2021, the European Citizens’ Initiative (ECI) “End the Cage Age” called on the European Commission to propose legislation to prohibit the use of cages for a wide range of farm animals. The Commission now assesses the feasibility of working towards the proposed legislation expected in 2027. The poultry breeding companies will then need alternative solution to safely establish pedigree of their stocks. Electronic nests relying on RFID can be used to establish a link between the egg and the layer (Marx et al., 2002) but they remain to be perfectioned in each concerned species to deliver reliable data. In addition, they can only help to build the maternal pedigree. By contrast, the use of molecular markers is susceptible to bring a complete solution to the issue.
This study concerns various populations of ducks, encompassing distinct species with diverse characteristics. The common duck (Anas platyrhynchos), which includes the Pekin duck, is extensively utilized in Asia for meat and egg production. The Muscovy duck (Cairina moschata), indigenous to South America, is prized for its supposedly lean meat. According to Jiang et al (2021), the divergence between these two populations occured around 14 million years ago. Additionally, the mule duck, a hybrid derived from crossing a Muscovy drake with a Pekin female, accounts for over 90% of the production of foie gras, a flagship of French gastronomy. The duck is therefore a major poultry species, for which the development of genomic tools promises to be no easy task, as markers should exist in the two species, and show variability.
Indeed, a microsatellite panel had been developed for duck populations in France (Chapuis et al., 2010), and was deemed usable in various purebred and crossbred populations. However, this panel exhibited assignment rates to a unique parental pair too low to be routinely used at a large scale, mainly because markers revealed to be poorly polymorphic within the Pekin and Muscovy populations (Chapuis et al., 2010).
Here we present the setting of an efficient and affordable assignment panel that can be used to assign pedigree in populations of Muscovy and Pekin ducks, as well as their hybrids. To build a posteriori the pedigree in these populations, the KASPar technology was retained, as providing access to affordable small SNP arrays. We will present and discuss its performances to assign pedigree in a genetic experimental design. The possible use of the developed molecular tools in other populations, such as local breeds, will also be discussed.
Material and methods
Designing the Assignation Panel
Development Strategy of a Cost-Efficient panel
As an important preliminary note, it is crucial to emphasize that our objective was to develop an assignment panel, not a set of markers for linkage analyses. The desired properties of these markers differ significantly. Specifically, the SNPs in the assignment panel are preferentially situated in “neutral” loci—regions where allele frequencies are not expected to be strongly influenced by selection, as might occur if the SNP were located near a QTL. We seek SNPs with high minor allele frequency (MAF) that segregate independently to maximize the number of possible genotype combinations, thereby enhancing the ability to discriminate between parental pairs. Namely, our objective was to assign pedigree in an experimental population of hybrid mule ducks and their purebred half-sibs, namely Muscovy duck (Cairina moschata) for the sire line and Pekin duck (Anas platyrhynchos) for the dam line. Therefore, we aimed at organizing mating plans and building an affordable 96 SNP panel to retrieve the pedigree using molecular information. The two parental lines pertained to populations sampled to previously develop the ThermoFisher Axiom HD SNP duck array, hereinafter referred to as 600K array (Teissier et al., 2019). This collection of genotypes, already available (hereinafter labelled as “reference dataset”), was used as a starting point to build the desired panel. The 600K genotypes from Anas platyrhynchos (n=139), Cairina moschata (n=79) and some mule ducks (n=45) were used to assess allele frequencies. However, as among these genotypes only 15% originated from the same populations as our parental lines, a two-step strategy was adopted. In a first instance, a set of 192 SNPs eligible for the chosen technology was developed, based on both their frequencies in the three populations and their technical properties. This first set was used to obtain first genotypes in our parental lines and in some triplets of mule progeny and their parents, i.e. with known pedigree. In a second step, the 96 SNPs with best technical outcomes and frequencies within and across parental lines were selected among these 192 to obtain an efficient panel. They were later used to establish pedigree of our offspring batches. Note that the mule duck is the hybrid obtained by crossing Muscovy drakes and Pekin females, while the common duck populations (Anas platyrhynchos) represented on the 600K reference dataset encompassed many other breeds than Pekin.
Selection of 192 SNP eligible for KASPar technology
The KASPar fluorescence genotyping technology was selected. In contrast to the AXIOM microarray-based technology, which is appropriate for genotyping a very large number of markers, KASPar is cost-effective for a smaller number of SNPs. Additionally, KASPar offers flexibility, allowing the genotyping panel to be adapted as needed. This adaptability will be particularly beneficial for easily switching from 192 to 96 SNPs. A diagram comparing the two technologies is provided in the supplementary figure S1. The 600K chip contained 334,950 SNPs segregating in the Muscovy duck library and 331,241 SNPs segregating in the common duck library. A preliminary step was to select only markers without polymorphism in the 50 bp before and after the SNP, as primer length is longer (50 bp) with the KASPar technology than with the Axiom technology (35 bp). For that purpose, pool-sequenced DNA from 50 males, sampled from several French populations (wild mallard and commercial Pekin and Muscovy) were used (Teissier et al., 2019). Primers for markers found on the 600K chip were aligned on the reference genome (Anas platyrhynchos genome from (Huang et al., 2013), and Cairina moschata genome from (Thébault et al., 2019)). Only SNPs exhibiting an identical primer sequence in the Muscovy and common duck populations were kept. After this step, 229,138 SNP remained in the Muscovy library while the common duck library contained 198,091 markers. The intersection of both led to a list of 399 candidate SNPs, susceptible to be amplified in the mule duck population. Only 396 were awarded the recommended PolyHighResolution status from the Axiom Analysis Suite software distributed by ThermoFisher, meaning they were found high quality and polymorphic. The final list of 192 SNPs was to be built among these 396, applying filters to individuals and triplet genotypes available in the reference dataset. PLINK V2.0 (Purcell et al., 2007) was used to perform filtering operations on missingness, both for genotypes and SNPs, minor allele frequency (MAF), and Mendelian mismatches. The retained criteria were values of 0.95 for call rate (CR) and call frequency, and 0.10 for MAF within Pekin and Muscovy populations. About 100 trios representing various genetic types were available in the reference dataset and could, therefore, be used to track markers leading to Mendelian incompatibilities. Such incompatibilities disqualified the concerned markers. An ultimate filter was applied based on linkage disequilibrium (LD), aiming to choose independent markers.
Setup of the final cost-efficient 96 SNP panel
A mixture of two groups of animals was used to evaluate the properties of the 192 selected SNP. The first group was a subset of the reference dataset composed of 72 individuals: 44 Pekin, 15 Muscovy and 13 mule ducks, in order to ensure consistency between KASPar and Axiom results. The second group encompassed most of the parents (133 Pekin and 127 Muscovy ducks) of the experimental batches to assign. To select the final 96 markers with desired properties, similar criteria as for the previous step were used: markers were kept when they had maximum call-rate of 5% missingness, a within line MAF of 0.15 and absence of Mendelian incompatibilities, the latter being assessed using samples with known kinship (nine offspring-sire-dam triplets in Pekin, four offspring-sire pairs and two offspring-dam pairs for mule ducks). The 96 selected markers were then combined on a single plate to genotype the offspring for reassignment.
Assessment of the assignment power of the 96 SNP panel
An evaluation of the assignment power of the marker set was carried out by computing the exclusion probability (Vandeputte, 2012), which is the probability of a randomly chosen parent-pair being genetically excluded as parents of a randomly chosen offspring, when that parent pair did not produce that offspring (Dodds et al., 1996). It depends on the number of parents and the allele frequencies in the parental population. It provides a good quality criterion for the set of markers once the parental population is genotyped.
Sample Collection and Genotyping
Blood samples from offspring and their parents were collected after slaughtering and sent to the INRAE genotyping platform Gentyane (Clermont-Ferrand, France) for DNA extraction and genotyping. Genomic DNA extraction was performed using GenFind V2™ (Beckman Coulter) commercial kit. The offspring were genotyped for parentage assignment using 96 SNP in KASPar . Dynamic Array™ IFC 96 * 96 chips were used with Biomark™ HD Reader to perform the competitive PCR and chip reading. The Fluidigm® SNP Genotyping Analysis software was used to analyze the genotyping results.
Parentage Assignment Validation in an Experimental Design
Ethical statement
The present study was conducted in agreement with the 2010/63/EU regulation for use of animals for research purposes. Animals were bred at the INRAE Duck farm (UEPFG, Benquet, France) which has been approved for animal experimentation (C40-037-1). Experiments were carried out following a protocol approved by the French Ministry of Higher Education, Research and Innovation, abiding by European regulations for animal care (APAFIS# 2018013116519672).
Mating design
The mating plan was designed with the double purpose of achieving pedigree assignment in a limited size population (our testing capacity did not exceed 280 ducklings in Pekin, 220 animals in Muscovy and mule ducks) with related breeders, while preserving enough genetic diversity in the offspring population to estimate genetic parameters. The retained strategy was i) to split related breeders in separate factorial designs and ii) to ensure that the largest possible number of maternal origins was represented among ducklings. Each female stock (N= 96 for Muscovy ducks and N=99 for Pekin ducks) was split in three 35 m² cells with slatted floor. These cells were equipped with nests lined with wood shavings to limit the number of floor eggs. To respond to the species specificities, 15 partially closed nests were available in each cell for the Muscovy ducks, whereas for the Pekin ducks, cells were equipped with two large collective nests without roof. Drakes (N=48 for Muscovy ducks and N=34 for Pekin ducks) were kept in individual cages, to avoid aggressive behaviors. A factorial design was implemented, in which groups of females within a given cell were inseminated with pre-designed semen pools from four drakes in Muscovy. In Pekin, the number of drakes per semen pool varied between three and four. In the Muscovy population, females from each cell were divided into four groups of eight individuals, whereas in the Pekin population, they were divided into three groups of eleven or twelve individuals. Each group was identified using a colored leg ring. Thus, in the Muscovy population, the number of possible parental pairs of an egg reduced from 48 males*96 females = 4608 to 3 cells*4 groups*8 females*4 males = 384. In the Pekin population, on the basis of the mating plan, this number has been reduced from 3366 to 375. Based on preliminary genotyping results, the maximum number of parental pairs in both populations was deemed sufficient for accurately estimating genetic parameters. Subsequently, dams and sires were assigned to each cell and grouped according to their relatedness, ensuring that siblings were not placed in the same group to avoid complications arising from their similar genotypes, which could hinder the performance of relatedness assignment software. During the two-week reproduction period, each group of females was repeatedly inseminated with pooled semen from the same group of drakes. Following common practices, insemination doses were calibrated to provide 100 million spermatozoids for Muscovy females and 150 million spermatozoids for Pekin females. Contribution of each male was monitored prior to mixing based on optical density of ejaculates, to provide an equal number of spermatozoids from each drake within an insemination dose.
Egg collection and hatching
Eggs were harvested daily during the egg collection period. Day of lay and cell number were written on the shell. After candling prior to the hatcher transfer, eggs were put into hatching baskets (one hatching basket per day of lay and cell number) and then were ordered in the hatcher based on decreasing number of viable eggs. At hatch, ducklings were identified with a wing band until the desired number of ducks was reached, i.e. not all hatching baskets were collected. Given the above-mentioned limited testing capacity and assuming a female lays only one egg each day, the ranking of the baskets based on egg numbers was retained to maximize the number of dams contributing to the final retained population. The correspondence between the wing band and the cell number was recorded.
A posteriori pedigree assignment
The experimental population to assign was composed of three batches, each related to a genetic type: 157 male Muscovy ducks, 207 male mule ducks and 273 Pekin ducks of both sexes, all issued from the parents first genotyped with the 192 SNP panel. The APIS software (Griot et al., 2020) was used for pedigree assignment. The two available methods were compared. One is based on the maximization of the average Mendelian transmission probability of the markers for a given offspring and all the possible parental pairs. The other one is based on the exclusion principle, where any Mendelian incompatibility eliminates a parental pair until only the true one remains. In order to account for genotyping errors, a user-tuned number of mismatches can be allowed and was set to two. Offspring exhibiting more than 5% missingness in genotypes were excluded from the assignment process, leading to the removal of 9 individuals (i.e. 3.3 % of the initial 273 offspring to be assigned) in the Pekin population only. Knowing the effective factorial design, we were able to produce a positive list of possible parental pairs and challenge the putative pedigree produced by the software with factual elements.
Results and discussion
First List of 192 Markers
Among the birds with 600K genotypes available in the reference dataset, only those exhibiting a call rate over 0.95 (i.e. with less than 5% of missing information) were retained, leading to a subset of 139 Pekin, 79 Muscovy and 39 mule ducks with genotypes, and a final number of 94 offspring sire dam triplets. Call-rate filtering for markers (maximum 5% missingness) led to a list of 348 SNPs, among which twelve were discarded because of Mendelian mismatch occurrences. SNPs were kept when minor allele frequency exceeded 0.10 in each of the Anas platyrhynchos and Cairina moschata populations, which led to a list of 232 SNPs. Only SNPs showing some polymorphism in the 39 mule duck samples were kept, reducing the number to 210. This criterion was applied to make sure the retained markers were not monomorphic among mules, as assignment of mule ducks was of prime interest. Finally, the list of 192 primers was obtained after eliminating SNPs exhibiting a LD above 0.25 with other markers.
Design of an Operational 96 SNP Panel
Elementary statistics about CR and MAF of the 192 SNP for our parental populations are displayed in table 1.
Table 1 - Call-Rate and Minor Allele Frequency (MAF) observed for the 192 SNPs in the parental populations
Anas platyrynchos N=133 | Cairina moschata N=127 | |||
|---|---|---|---|---|
Call-rate | MAF | Call-rate | MAF | |
minimum | 0.940 | 0.026 | 0.258 | 0.047 |
1st quartile | 0.993 | 0.222 | 0.984 | 0.236 |
median | 0.993 | 0.338 | 0.992 | 0.323 |
3rd quartile | 0.993 | 0.412 | 1.000 | 0.418 |
maximum | 1.00 | 0.500 | 1.000 | 0.500 |
These results were obtained for the parents of our experimental populations (133 Pekin and 127 Muscovy ducks), which explains why MAF were lower than 0.1 for some markers, as initial thresholds were set on a different population (our reference dataset). In our experimental Muscovy population call-rates were lower than expected. Fifty-seven SNPs exhibited missingness rate ranging from 0.42 to 0.75, while they were below 5% in the Muscovy samples previously genotyped with the 600K chip. Our hypothesis is that undetected polymorphisms in the primer sequences can be incriminated for these poor results. Such polymorphisms remained undetected in the few individuals sampled from the same line as our experimental populations. These 57 SNPs were discarded from the list. This endorses the strategy of starting with 192 SNPs to retain a final list of 96. Six additional markers exhibiting at least one Mendelian mismatch were deleted, reducing the list to 133. The minimal MAF criterion was set to 0.10 in each parental population, resulting in a list of 111 SNPs. Finally, to ensure desirable properties in the mule duck population, 7 SNPs with a call rate below 0.95 in the 39 mule samples were discarded. Eight additional markers were thrown away based on the clustering quality of their genotypes in the Fluidigm® SNP Genotyping Analysis software, resulting in the final list of 96 SNPs.
The final list of 96 SNP is displayed on table 2, while the list of 192 markers including the MAF in the three populations represented in the reference dataset is given as supplementary material. The mean and median distances between SNPs were 2,238,171 bp and 4,433,066 bp, respectively, in the 192 SNP panel, and 3,919,876 bp and 7,545,704 bp, respectively, in the 96 SNP panel. These data suggest that the SNPs are widely dispersed across the genome, a characteristic anticipated for markers used in an assignment panel.
The MAF distributions of these 96 SNP in our experimental populations are displayed on figure 1. Figure 2 shows the location of the SNPs on the different chromosomes. The localization of the 192 SNPs (upper panel) was somehow consistent with the size of chromosomes, with a larger number of SNPs on macro-chromosomes compared with micro-chromosomes. No SNP was located on chromosome 17 and 23. For the final set (lower panel), the priority was given to technical proprieties of the markers, leading to some gaps (no SNP on chromosomes 13, 15, 17 and beyond 25) and only two on chromosome 1. Nonetheless, the vast majority of SNPs in the final set were located on macro-chromosomes (numbered from 1 to 8, following (Skinner et al., 2009)). Supplementary Figure S2 illustrates the distribution of SNPs on the Axiom HD chip and the 399 SNPs shared between the Anas platyrhynchos and Cairina moschata datasets. Notably, chromosome 17 is absent from the Axiom chip. Twelve of the 399 common SNPs were located on chromosome 16, which may explain the relatively large representation of chromosome 16 in the final panel of 96 selected markers.
A consistency (i.e. percentage of identical genotypes) of 0.997 was observed between the genotypes of the 72 individuals in the reference panel, which were obtained with both technologies (KASPar and Axiom). Three individuals were genotyped twice with the KASPar technology with complete (100%) consistency. As previously stated, the set of animals used to obtain the final list of 96 markers contained nine individuals of complete known pedigree. Using these 96 SNPs, all offspring in the 9 trios of known pedigree were correctly assigned to their true parental pair using the APIS R package. All these factors reinforce our confidence in the panel’s effectiveness for reassignment in target populations.
Table 2 - Name and position of the 96 SNP retained in the final list.
Position refers to the Anas platyrhynchos library. KB745320.1 is a scaffold.
Chromosome | Position (bp) | Marker name | Chromosome | Position (bp) | Marker name |
|---|---|---|---|---|---|
1 | 109061561 | AX-247363485 | 7 | 639397 | AX-247355830 |
1 | 198136954 | AX-247363213 | 7 | 6642882 | AX-247355836 |
2 | 9314971 | AX-247354978 | 7 | 6784807 | AX-247364551 |
2 | 22038866 | AX-247363748 | 7 | 7458603 | AX-247364557 |
2 | 25524298 | AX-247355025 | 7 | 7903291 | AX-247355848 |
2 | 48224427 | AX-247355091 | 7 | 17149047 | AX-247364577 |
2 | 57105300 | AX-247363838 | 7 | 37659499 | AX-223686578 |
2 | 72878000 | AX-247363840 | 8 | 5024747 | AX-247355910 |
2 | 95527796 | AX-247355149 | 8 | 9828535 | AX-247364640 |
2 | 106227402 | AX-247363883 | 8 | 18077068 | AX-247355936 |
2 | 125817433 | AX-247355201 | 8 | 20064891 | AX-247364660 |
2 | 130944301 | AX-247363942 | 8 | 23941232 | AX-247364672 |
2 | 133449691 | AX-247363956 | 8 | 25365172 | AX-247364675 |
2 | 142558953 | AX-247355235 | 8 | 26073249 | AX-247364679 |
2 | 148407413 | AX-247355249 | 9 | 6446865 | AX-247364711 |
2 | 152370825 | AX-247355261 | 9 | 10829712 | AX-247356029 |
2 | 152906965 | AX-247355267 | 9 | 11668820 | AX-247364749 |
3 | 178108 | AX-247364000 | 9 | 13906991 | AX-247364763 |
3 | 22898020 | AX-247355316 | 9 | 14469818 | AX-247364765 |
3 | 34203102 | AX-247364053 | 10 | 11096372 | AX-247356148 |
3 | 41332352 | AX-247364072 | 11 | 15392465 | AX-247364917 |
3 | 49962556 | AX-247364080 | 12 | 4812384 | AX-247356238 |
3 | 53539930 | AX-247355356 | 14 | 6336270 | AX-247356370 |
3 | 66856580 | AX-247364116 | 14 | 14544447 | AX-247365129 |
3 | 68837303 | AX-247364118 | 14 | 14827130 | AX-247365133 |
3 | 74410901 | AX-247364122 | 16 | 2984766 | AX-247356455 |
3 | 110150507 | AX-247355450 | 16 | 3718731 | AX-247356463 |
3 | 110627101 | AX-247355452 | 16 | 9044628 | AX-247356481 |
4 | 6220620 | AX-247364191 | 16 | 9063242 | AX-247356483 |
4 | 14309946 | AX-247355482 | 16 | 13744076 | AX-247365233 |
4 | 25865050 | AX-247355506 | 16 | 14448873 | AX-247356512 |
4 | 60721998 | AX-247364276 | 18 | 5084874 | AX-247356525 |
5 | 2505593 | AX-247364303 | 19 | 10473301 | AX-247365299 |
5 | 6739459 | AX-247364317 | 19 | 10494308 | AX-247365301 |
5 | 7253477 | AX-247364320 | 20 | 2186582 | AX-247365309 |
5 | 26939905 | AX-247364353 | 20 | 6341095 | AX-247365329 |
5 | 27529421 | AX-247355645 | 20 | 8185628 | AX-247356614 |
5 | 36094216 | AX-247364375 | 20 | 9156425 | AX-247356617 |
5 | 42690814 | AX-247355671 | 20 | 11133474 | AX-247365344 |
5 | 45865637 | AX-247355673 | 21 | 12160575 | AX-247365370 |
5 | 54586500 | AX-247355693 | 22 | 2485025 | AX-247365384 |
5 | 54717023 | AX-247364410 | 22 | 2576730 | AX-247365386 |
5 | 58368151 | AX-247364419 | 24 | 2923576 | AX-247365450 |
5 | 58440894 | AX-247364421 | 24 | 4729515 | AX-247365466 |
5 | 59563000 | AX-247355708 | 24 | 5386920 | AX-247356749 |
5 | 62080514 | AX-247355726 | 25 | 1070592 | AX-247356762 |
6 | 28171519 | AX-247364508 | 25 | 1255481 | AX-247356766 |
6 | 31606612 | AX-247364526 | KB745320.1 | 252400 | AX-247364465 |
Following Vandeputte (2012), the exclusion power of the 96 SNP panel, based on the allele frequencies in the parental population and assuming random mating, was computed and found above 0.99999 in all the populations. These values were an encouraging result before attempting to build the pedigree of our experimental batches.
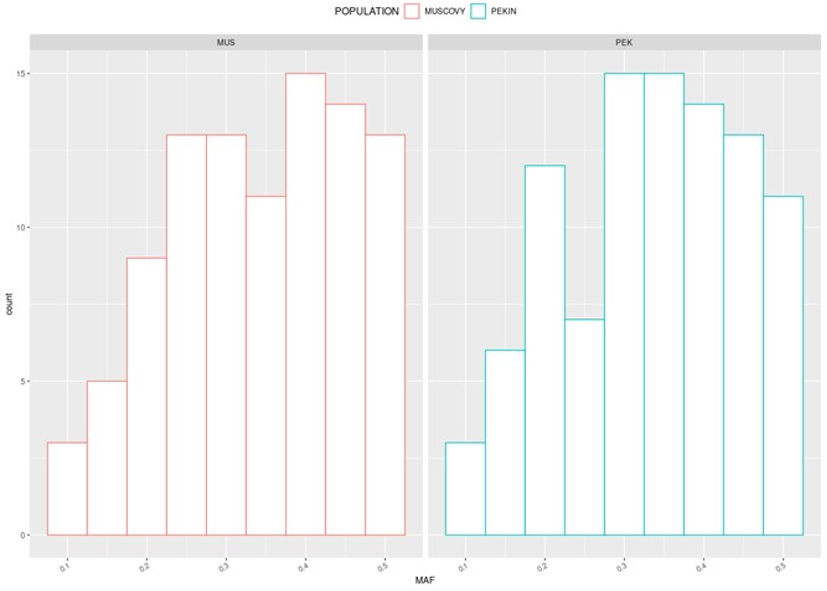
Figure 1 - Minor Allele Frequency distribution of the final 96 assignment markers in the experimental population
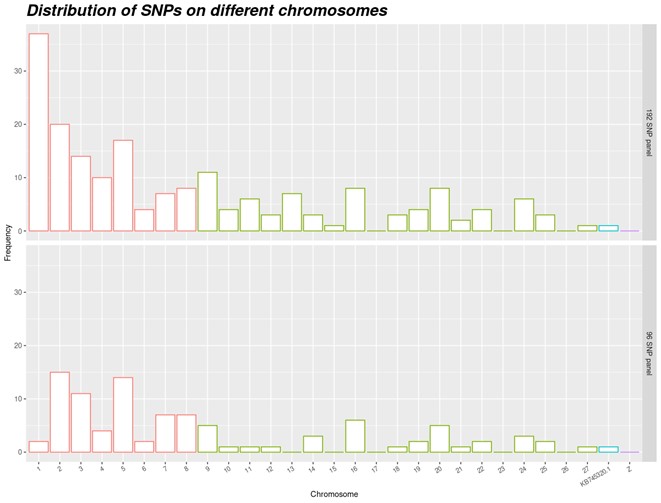
Figure 2 - Location of the SNPs on the chromosomes (upper part: 192 SNP panel and lower part: 96 SNP panel). Chromosomes 1 to 8 are macro-chromosomes, chromosomes 9 to 27 are micro chromosomes, Z is a sexual chromosome and KB745320.1 is a scaffold.
Obtention of a DNA-based Pedigree of our three Experimental Populations
Assignation rate
The assignment rate to a unique parental pair was 97% for the mule ducks (201 over 207), 95% in the Pekin population (260 over 273), and 87% in the Muscovy population (136 over 157). A posteriori, this poor performance in the Muscovy population could be attributed to the absence of 16 parental samples in the genotyped populations (fourteen females and two males). Yet, with 87% of success this set of markers performed at least as well as the previous microsatellite panel (Chapuis et al., 2010). In this study, assignment failures occurred when the most probable putative parent pairs identified had a relatively high number of Mendelian incompatibilities (above eight, when the threshold was set to two mismatches). In addition, this was confirmed by the two-peaked distribution of the difference in Mendelian transmission probability between best and second-best putative parents (figure 3) for the Muscovy offspring, unlike the two other populations. As stated by Griot et al., (2020) assuming a sufficient power of the panel (exceeding 0.99999 here), this situation clearly signaled missing parents. This demonstrates that the main obstacle for a posteriori building of pedigree is the absence of one or both parents. To confirm this hypothesis, the absence of the same number of parents (two sires and fourteen dams randomly discarded) was simulated in the Pekin population and, over 50 replicates, the average assignment rate dropped to 0.80±0.01, i.e. a loss of 16 percentage points. In these replicates, the maximum number of observed mismatches in the assigned individuals was 2, while, in the non-assigned Muscovy individuals, it ranged between 5 and 11, indicating a clear cut-off when one parent is missing. Another cause of APIS assignment failures may be the wrong estimation of the empirical threshold to be set in Mendelian transmission probability. According to (Griot et al., 2020), a minimal number of 200 offspring is required to properly estimate this threshold, while we had only 157 Muscovy.
Benefits of Mating Plan Knowledge
The outcomes of an APIS run can be split into three situations: i) direct successful assignment to the rightful parental pair, ii) wrong assignment to an erroneous parental pair, or iii) failure to return a unique parental pair. In our case, thanks to the availability of the mating plan, the two latter situations could be sorted out in most cases. As an illustration, the vast majority of assigned parental pairs was fully compatible with both the list of possible mating and the cell number where the egg was collected, associated with the wing band. They were, therefore, considered as correct, and corresponded to case i. In addition, these pieces of information allowed to detect and fix one single wrong assignment returned by the software. In this case, the parental pair ranking first on Mendelian transmission probability could materially not be the true one, unlike the second ranking pair, exhibiting a Mendelian transmission probability only slightly lower than the first one (case ii). Assignment failure (case iii) occurred in very few situations (less than 5% of cases in Pekin and mule populations), for instance, when the two most probable parental pairs featured the same sire while the different dams could not be separated based on Mendelian transmission probability only. In these cases also, supplementary information brought by the wing band, which identified which cell number the egg originated from, and thus which mating was possible, helped to designate the true pair among the putative pairs proposed by APIS.
Consequences on the Population Structure
Avian pedigreed populations are usually bred using individual cages for females, applying a hierarchical mating design (a single male used to inseminate p females, a dam having offspring from one sire only). In factorial designs allowed by group housing, a female can give birth to ducklings with multiple sires, up to four different drakes in our case. Table 3 displays, for each of three experimental batches, the proportion of dams which had progeny identified from k males, k varying from 1 to 4. The population structure here is different from a hierarchical mating design, as less than half of the dams had offspring from only one sire. This remarkable change in the mating design is displayed on figure 4, which shows the last batch of Pekin and its two generations of closest ancestors (parents and grand-parents). When the hierarchical mating plans operated, much less combinations of sires and dams were recruited than when the mating scheme was factorial. Population structure varied among the three genetic types displayed in table 3. Without any replicate, however, it is not possible to infer the differential consequences to be expected in the three populations once the hierarchical mating plan is replaced by a factorial one.
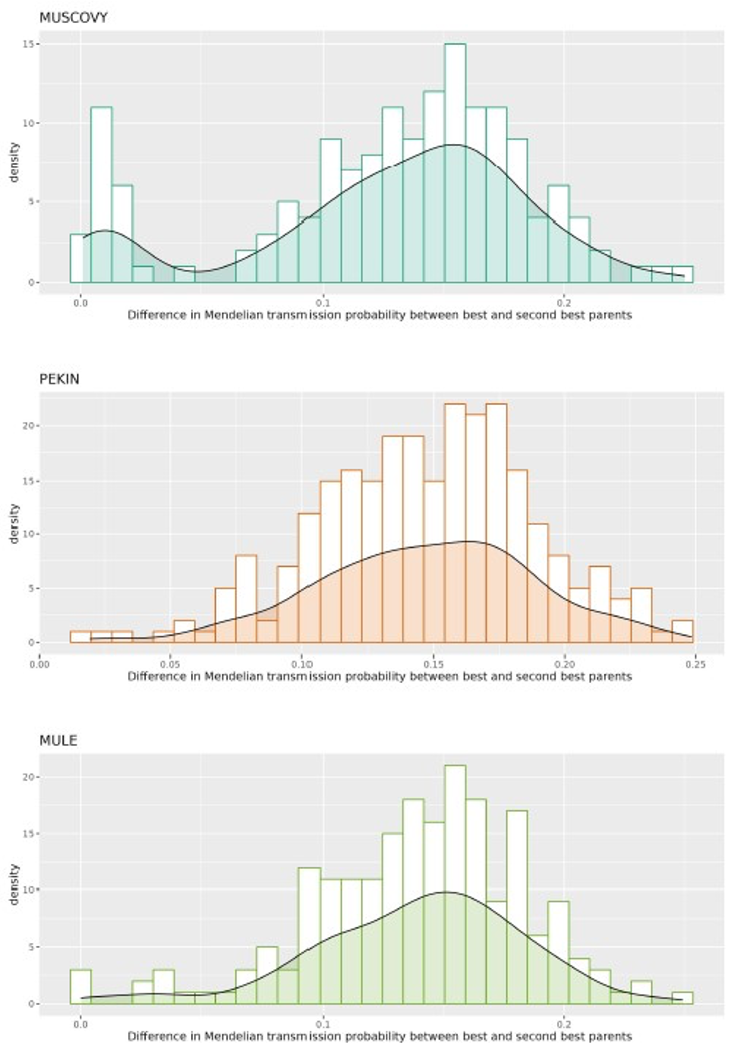
Figure 3 - Distribution of differences in Mendelian transmission probability between best and second-best putative parents in the three populations
It is useful here to remind that, given the characteristics of poultry reproduction, in particular the presence of sperm storage tubules in the oviduct of females, the hierarchical mating plan carried out for a long enough period was, regardless of the housing system, the only way to ascertain the pedigree of newborn chicks before the availability of molecular tools allowing for parentage assignment. Thus, females could be housed in cages or in pens, but they were mated to a single male during a given egg collection period. Switching from hierarchical to factorial design is recommended first for practical convenience when individual cages are banned: it is indeed easier to pick a female based on its colored leg ring and inseminate it with a prepared semen pool than randomly pick a female, read its wing band and inseminate it with sperm from the single relevant male. Besides, not only the SNP panel allows for parentage assignment but it also provides context for the correct estimation of maternal effects, which are no longer confounded with a sire-dam interaction in a given laying series, as can be seen on figure 4.
Table 3 - Proportion of dams giving birth to ducklings with k different sires
Population | |||
|---|---|---|---|
k | Cairina moschata | Mule ducks | Anas platyrynchos |
4 | _ | 33% | 12% |
3 | 16% | 41% | 30% |
2 | 36% | 20% | 27% |
1 | 48% | 6% | 30% |

Figure 4 - Pedigree representation of the last two generations in the Pekin line. Orange circles represent dams and blue circles sires. The upper part describes a hierarchical design (only one line originates from each orange circle, as each dam is mated with only one drake), while a factorial design is used in the lower part. In that case, females can have progeny with up to 4 males.
In the context of duck breeding for fat liver production, such a change in breeding schemes is prone to dramatically impact the way Pekin lines (i.e. the dam pathway of the mule duck) are selected. Indeed, their breeding values used to be computed based on purebred performances (body weight and laying performances) and crossbred performance measured on mule offspring. When these offspring are obtained through a hierarchical mating design, the dam estimated breeding value is confounded with the Muscovy drake potential, which may lead to bias, if the sire breeding value is not properly estimated, a common situation when evaluations for both Pekin and Muscovy lines are not carried out simultaneously. If mule offspring are obtained with multiple drakes for each dam, the bias partly wipes out. Besides, in the case of low male fertility, a Pekin female will potentially have lesser progeny with a hierarchical mating design than with a factorial mating design, due to the male side. Switching from a hierarchical to factorial mating design should, therefore, improve the selection process on the dam pathway.
Table 4 - Dam family structure in three successive batches of mule ducks. M1 and M2 were obtained using individual cages and a hierarchical mating design. M3 was obtained using a factorial design and pedigreed through genotyping.
batch | # anim | # dam | Dam family size | MEAN | VAR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||||
M1 | 247 | 87 | 4 | 17 | 55 | 11 |
|
|
|
|
| 2,84 | 0,49 |
M2 | 282 | 84 | 10 | 15 | 22 | 18 | 14 | 2 | 2 | 1 |
| 3,36 | 2,33 |
M3 | 204 | 69 | 18 | 14 | 15 | 11 | 4 | 2 | 4 |
| 1 | 2,96 | 3,40 |
On the other hand, management of breeding resources raises new issues in the case of floor reproduction and late pedigree knowledge. When the parents of the egg are known at egg collection (i.e. with a hierarchical design applied to individually caged females), it is easy to monitor family size at hatch and obtain a balanced family representation for a given batch size. This can be assessed looking at table 4, which displays dam family structure in three successive batches of mule ducks. In the latest mule batch (obtained under factorial design), dams had from 1 to 9 offspring, with an average of 2.96 ±1.88. Only 70% of the dams had male offspring in this latest batch. This proportion was above 85% in the previous batches- with hierarchical designs. This drop can originate from the sampling of ducklings at hatch (males kept until the desired number was reached), when the dam is not yet known, and may also be due to zootechnical issues, if some females did not lay hatchable eggs, or only floor eggs. Such an unbalanced contribution of breeders to the progeny due to free mating system has been described by Brard-Fudulea et al. (2023) in red partridge. Therefore, pen size (cell size in our situation) and animal sorting should be carefully organized, lest origins may be lost. In addition, there is room for optimization of the mating design. Usually mating plans are designed in order to monitor the increase of inbreeding rate, for instance by avoiding common ancestors between associated groups of males and females. Here another constraint should be imposed on the common ancestors within a group of breeders, lest difficulties arise to find the true parental pair. One solution could be to use, in the optimization process, a kinship matrix based on genotypes instead of the numerator relationship matrix derived from pedigree. One could also imagine minimize the expected inbreeding of future progeny, as do most mating plan setup software, while setting a constraint on a molecular kinship of breeders computed using marker genotypes. A similar algorithm (simulated annealing mixed with Lagrangian multiplier) was used by Chapuis et al. (2016) to optimize breeder selection under a constraint on kinship.
Last, but not least, here females were inseminated and doses were calibrated to equilibrate male contributions. Ultimately, in breeding companies with large populations, one could be tempted to rely on natural mating, using pens with p males and q females, like at the multiplication stage. Such condition would add another heterogeneity factor with mating behavior likely to dramatically impact family composition. A thorough modeling of selection schemes is, therefore, necessary, to face the replacement of hierarchical mating design with factorial ones.
Assignation power in other duck and poultry populations
This 96 SNP panel was explicitly designed to perform in our experimental population. Yet, eight 95 x 96 chips were used to obtain 192 SNP genotypes, leaving some spots available that were used to collect genotypes for local breed samples. Thirty-four Duclair and 10 Rouen individuals (two local breeds of Anas platyrhynchos) were thus genotyped. Minor allele frequencies averaged 0.26 and 0.29, respectively, in these two populations. These values were lower than those reported for our experimental lines in table 1. They nonetheless led to exclusion probabilities above 0.99 in these two populations, giving way to a potential use for improved management of genetic resources. Practically, a side outcome of this study is a list of 135 SNPs (i.e. the initial list of 192 SNPs, deprived of the 57 markers that did not work in our Muscovy population) with reliable properties being now available in Anas platyrhynchos, Cairina moschata and their hybrid offspring, to setup SNP sets for any commercial or local population. Commercial populations undergoing genomic selection are not concerned with the need of an efficient assignment marker set, as the thousands of SNP on a chip can also be used to build pedigree. Yet, the question remains for mule offspring, as usually, for cost reasons, only selection candidates are genotyped using medium or low density (MD) chips featuring 10 to about 50K SNPs. Should individual cages be banned in European breeding companies, mule ducks would also require genotyping and then the cost benefit ratio of using a 96 SNP set vs. a MD chip should be carefully reevaluated.
As previously stated, the setup of an operative assignment panel is not an issue in widely distributed poultry species, where genomic material has already been developed (chicken, turkey, ducks). This can be more complicated with minor species such as guinea fowl or game (partridge or pheasant). Yet, in Europe, breeders operating in these species could be also concerned with the ban of individual cages. Recently, in red partridge, assignment rate reached 90% using a 96 SNP panel (Brard-Fudulea et al., 2023). In their review, Flanagan and Jones (2019) noted that as few as 31 SNPs could be used to assign all offspring with >99% confidence in a population of wild birds. They also reported many examples (mostly in fish, some in mammals) where 96 SNP panels would be sufficient to provide a unique parental pair for each offspring. In our situation, we benefited from previous work carried out in ducks and the availability of a 600K microarray. Assignment panels could also be obtained de novo using Next Generation Sequencing (NGS) methods. As stated by Guichoux et al. (2011) these technologies enable the identification of large numbers of microsatellite loci at reduced cost in non-model species. Consequently, more stringent selection of loci is possible, thus further enhancing multiplex quality and efficiency. This potentially could allow for a microsatellite panel avoiding the pitfall encountered by Chapuis et al. (2010) where the available microsatellites were not sufficiently polymorphic in both parental populations simultaneously. NGS methods also provide different ways to obtain sets of SNPs that could be used for parental assignment. For instance, in Atlantic salmon, Holman et al. (2017) used RAD markers (Miller et al., 2007) to identify SNPs to be developed into a marker set. Knowledge of the mating plan allowed for a 100% accuracy in parentage resolution with no more than 94 SNPs, even when putative parents were related. These results, in accordance with our own, leave to hope that a set of 96 SNP and some practical rules for bird management could be enough to provide an affordable tool for effective parentage assignment in most commercial poultry populations.
Conclusion
In this study, starting from a 600K AXIOM chip, a 96 SNP panel was developed and proved effective for correctly assigning parentage in an experimental population of three connected genetic types. The technical process, including an intermediate selection of 192 SNPs evaluated in the populations of interest, highlighted the importance of careful marker selection when transferring between technologies (AXIOM to KASPar). Besides, as poultry populations have limited effective sizes, an optimization of the factorial design was needed to avoid genetically similar types of progenies in the same pen (issued from sibling breeders), which resolved most of the dubious assignations, and the pending ones actually pointed out missing samples in the parents.
If the 96 markers in the final panel were the best suited for our objective, the 192 SNPs in the intermediate panel (or rather 135, as 57 were problematic in our Muscovy population) can be used to develop panels that offer the highest reassignment rates, depending on the populations (commercial or heritage) being reassigned.
The ban of individual cages is likely to dramatically impact selection schemes in poultry species. Here, we suggest to switch from a hierarchical to a factorial mating design, which leads to clear changes in the population structure. Their consequences in the long term for selection schemes still remain to be investigated, and the management of the mating plans (i.e. pen size) will have to be optimized accordingly. In addition to an impact on the pedigree, banning individual cages will also affect the individual recording of laying traits, and the development of connected nesting devices to record laying performances of female ducks will also be a concern.
Appendices
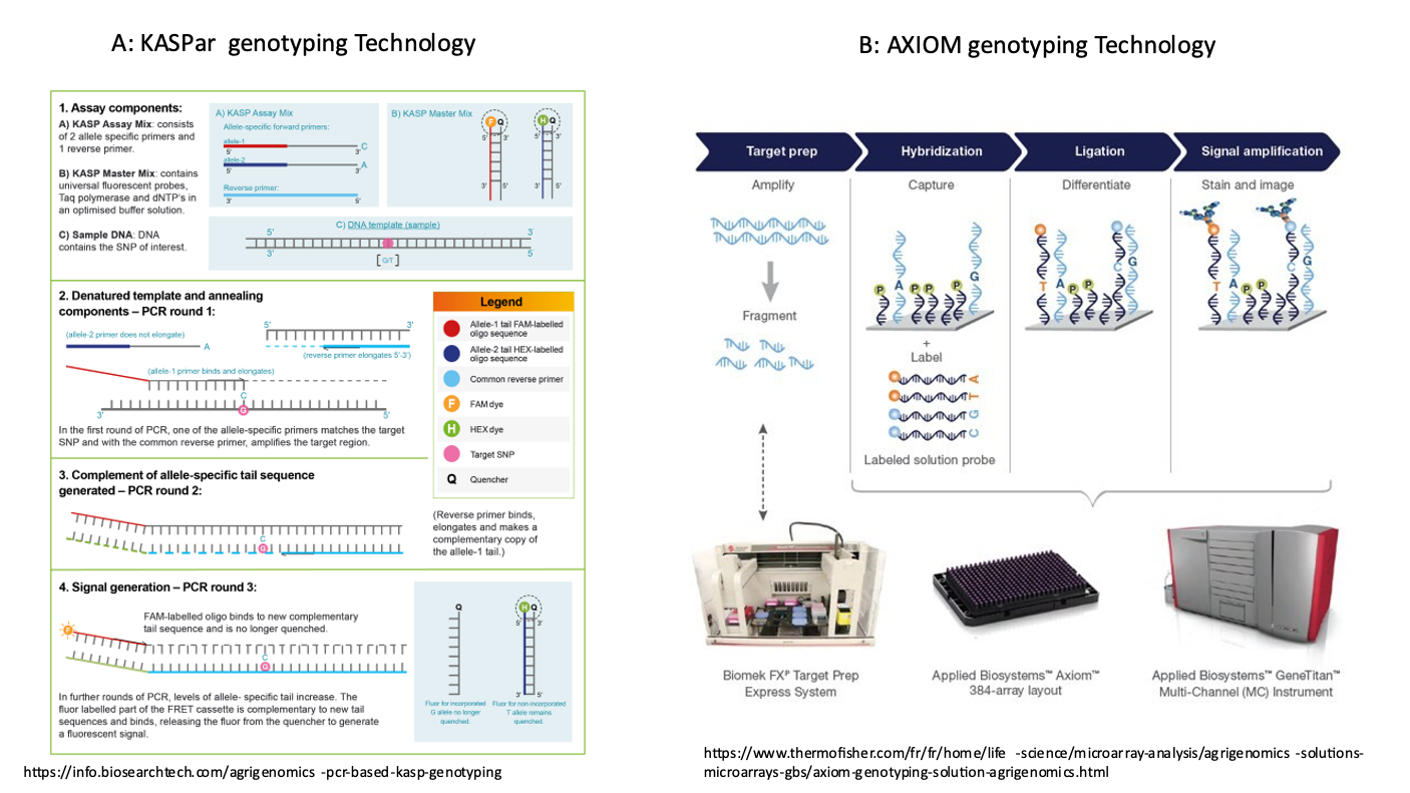
Supplementary figure S1: Differences between the two genotyping technologies described in the article.
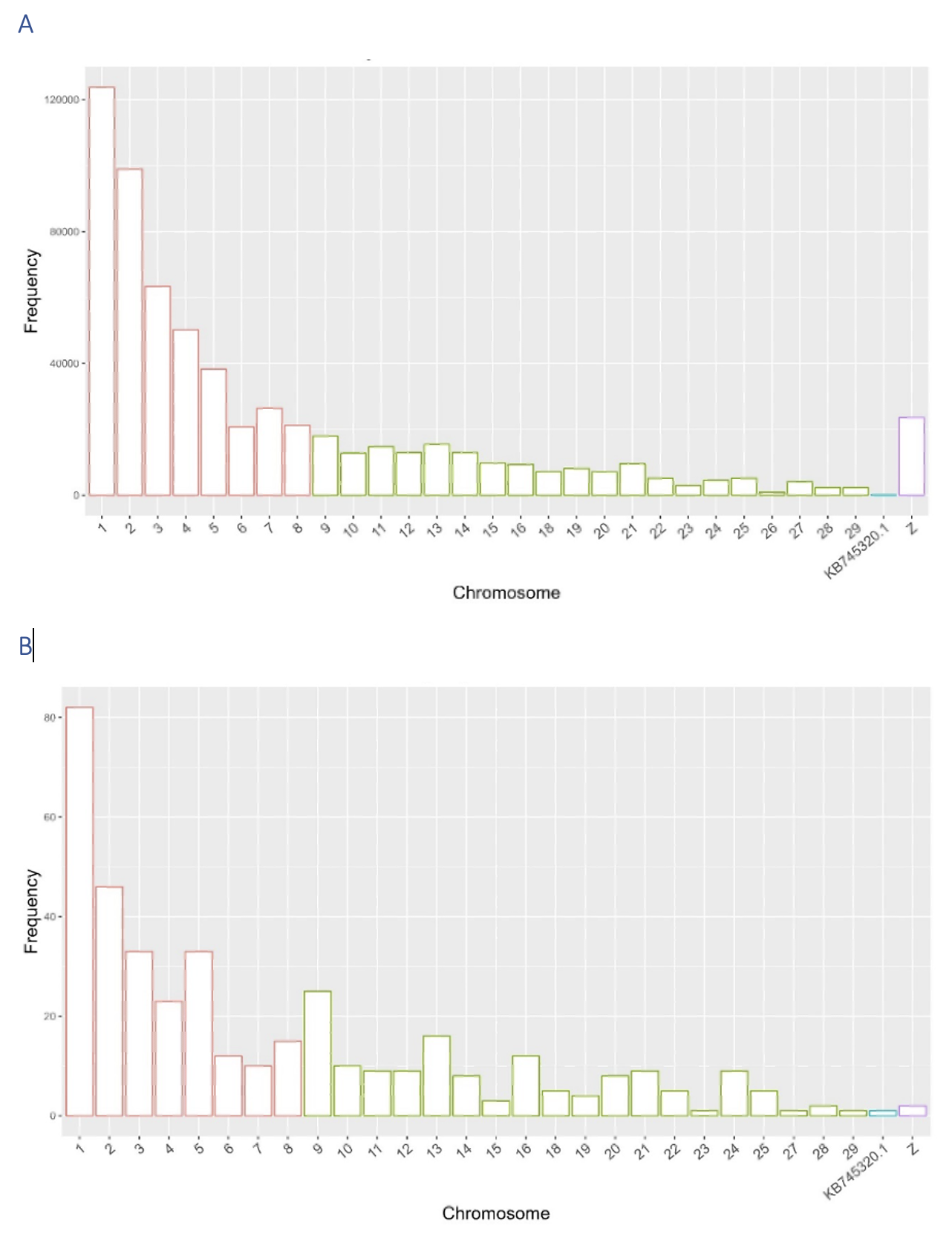
Supplementary figure S2: Distribution of SNPs across chromosomes: Panel A shows SNPs from the 600K chip, while panel B presents the 399 SNPs common to both Anas platyrhynchos and Cairina moschata libraries. Chromosomes 1 to 8 are macrochromosomes, 9 to 27 are microchromosomes, Z represents a sex chromosome, and KB745320.1 is a scaffold.
Acknowledgements
The authors are grateful to Grimaud Frères Sélection (Sèvremoine, France) for giving access to the results of 600K genotypes in several commercial populations of Pekin and Muscovy ducks. In addition, they sincerely thank INRAE-Gentyane (Clermont-Ferrand, France) for carrying out the genotyping. Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Animal Science (https://doi.org/10.24072/pci.animsci.100304; Rafat, 2024).
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
Data availability
Data (genotypes, putative pedigree and R script) are available at https://doi.org/10.57745/XQLL2U (Chapuis, 2024)
Funding
This work was supported by an INRAE Animal Genetics division grant (ASPARCAN program, 2021).