
 CC-BY 4.0
CC-BY 4.0
Introduction
The behaviour of an animal can inform about the internal state of that animal, in relation to biological functioning, health and welfare. In farm animals, activity measurements have long been used to identify differences in walking behaviour to detect lameness (Pastell et al., 2009) or differences in the amount of activity to detect oestrus (Saint-Dizier & Chastant-Maillard, 2012) or periparturient disorders (Rutten et al., 2017; Rutten et al., 2013; Weary et al., 2009). Additionally, comparing the activity of a focus animal to its baseline or to pen-mates allows identification of deviations that potentially indicate a change in internal state, e.g., an animal becoming ill (Kok et al., 2023). The continuous measurements of activity allow for complex patterns to be highlighted, such as circadian components and regularity, and these patterns can be used to identify animals at risk of diseases (van Dixhoorn et al., 2023; Van Dixhoorn et al., 2018; Wagner et al., 2021).
Ethologists, and other researchers usually calculate several metrics from activity data to characterise an animal’s behaviour, e.g., time spent on each activity, fragmentation of activity, 24-h patterns or proportion of animals engaged in the same activity. However, the way these metrics are calculated vary between studies. For instance, the synchrony between animals, that reflects the functioning of a group, can be calculated at individual level as the percentage of animals performing the same activity as the focus animal, then at group level as a mean of that percentage or using concordance indices such as Kappa coefficients or overdispersion index (Raussi et al., 2011; Tuomisto et al., 2019; Veissier et al., 1990). The pros and cons of each metric are rarely explained.
The activities can be documented by observers, from direct observation or from video recordings, or obtained using sensors. Direct observation allows precise activities to be recorded. For instance, lying can be divided into lying head down versus head up with corresponding arousal levels inferred (Veissier et al., 2001). Direct observation (both real time or from video recordings) creates a high workload and data at night are often missing due to difficulties in observing animals in the dark. The last ten years have seen a boom in the development of sensor technologies, which can provide data along time series more easily than direct observation. Activities (especially in large domestic animals like cattle) can now be recorded continuously on individual animals and for very long periods with little workload, using accelerometers, image analysis from videos, or Real Time Locating System (RTLS) (Buller et al., 2020). Most of the commercially available sensors that monitor cattle activity show excellent performance in validation studies (Lee & Seo, 2021). They usually provide information on gross activities such as lying, standing, moving, feeding, and ruminating or the position of animals in the barn. From the organisation of these activities, specific patterns can be detected, especially those indicative of animal malaise due to illness or stress (Wagner et al., 2021) or related to the social organization of animal groups (Rocha et al., 2020), allowing new insight into animal behaviour. However, to date the flood of data obtained from sensors seem under-utilised (Koltes et al., 2019).
Access to data on activity is not only facilitated for ethologists, but also for researchers from other discipline, e.g., animal health and animal production and for non-scientists like users of precision farming systems that are not necessarily used to process data on behaviour (Borchers et al., 2016). It is thus crucial to provide harmonised metrics to analyse behaviour and to discuss what they are supposed to measure and their limits. This would allow comparisons between studies, help the interpretation of results, extend the use of activity data beyond ethologists, and ease the re-use of datasets.
In the present paper, we consider the metrics that describe different aspects of behaviour from data collected by sensors or by direct observations (Figure 1). We consider metrics describing 1- overall activity, 2- fluctuations around mean activity, 3- cyclicity of activity, and 4- synchrony between animals. For each of these four groups of metrics, we identify the main metrics in use, the calculation methods, the limitations or the difficulties that can be encountered to calculate them, and their biological meaning (i.e. what it implies for animal welfare or health or the functioning of social groups). Examples are taken essentially from cattle, for who individual activity sensors are easily available at present. The calculations, however, can apply to datasets from other species (including other animal-specific activities) and can be performed on data coming from sensors as well as from observation. This paper does not focus on the validation of the sensors or the observers (to assess inter or intra observer variation) but on the processing of the data acquired by the sensors or observers.
A methodological exploration is pivotal and timely given the boom of activity data obtained by sensors. Our paper will hopefully support researchers by improving the use of activity data to answer their research questions. In addition, it should facilitate the consideration of animal behaviour by non-ethologists especially in Precision Livestock Farming (PLF), with a view to help phenotyping animals for selection, monitoring them for the detection of changes due to specific states such as oestrus, disease, or stress, or evaluating housing conditions and management aspects.

Figure 1 - Visualisation of the data acquisition and processing. The scope of this paper focusses on the possible calculations of the data that describe the four metrics: overall activity, fluctuations around the mean activity, cyclicity of activity and synchrony. The calculations can be done on data that is acquired by observations and/or with sensors.
The data
In the present paper, we focus on gross activities such as feeding behaviour (including eating and drinking), active behaviour (apart from eating or drinking) including walking, running or other movements, or inactive behaviour including standing still and lying. More specific activities include grazing and ruminating in ruminants (e.g., cattle, sheep, goats), rooting in pigs, and foraging behaviour in poultry. All these activities are characterised by lasting for some time. They are sometimes referred to as ‘states’, as opposed to brief behaviours (e.g., interactions between animals) that are referred to as ‘events’.
Observers can perform different types of sampling methods i.e., continuous, meaning they note changes in activity over a certain period of time and record the time the change occurred. Alternatively, observers can perform scan sampling i.e. they note the animal’s activity as detected at first glance at regular intervals (Bateson, 2021). Examples of the calculations with continuous and scan sampling data are shown in the supplementary materials. A mix of the two is also possible: the activity is recorded continuously and at the end of each interval (e.g. 5 min) the observer notes the predominant activity; the format of data will then be similar to that of scan sampling. Sensors generally produce a signal that is nearly continuous; the data on gross activity are usually delivered as time spent in each activity per time intervals (e.g. minutes per hour or per 15 min) or as predominant behaviour per time interval. The metrics that can be calculated, depend on the formats of data (see next sections).
Metrics to address overall activity
Definitions
The overall activity refers to the time spent performing specific activities during a certain time period. Each activity such as feeding, drinking, walking, standing idling or lying, can be characterised by the duration it is performed, the number of bouts (where a bout is defined by the continuous expression of an activity), and the average duration of bouts. The overall fractioning of activity refers to how many bouts of activity are noticed, in other words, how often the animal changes between activities. The level of activity reflects how much an animal is active, that is walking, running, or feeding rather than lying or standing immobile. The time period on which these metrics are calculated varies between studies, e.g. an hour or a day. The term ‘time budget’ is specifically used to describe how an animal divides its day (or shorter period) into the various activities.
Calculations
Proportion of time spent in an activity.
The proportion of time spent in a given activity a in a collection of activity bouts B (whatever the activity) is calculated as follows:
-
\({Proportion\ OfActivity}_{a}^{B} = \frac{\left( \sum_{b \in B,\ \ \ A_{b} = a\ }^{}D_{b} \right)}{\sum_{b \in B}^{}D_{b}}\)
where Db is the duration of the bout b and Ab is the activity in bout b.
In case of scan sampling, the time spent in an activity is estimated from the number of scans per activity, multiplied by the interval between scans - although what the animal has done between scans remains unknown. The proportion of time spent in an activity a in a period P can be calculated as follows:
-
\({Proportion\ OfActivity}_{a}^{P} = \frac{\left( \sum_{p \in P,\ \ A_{p} = a\ }^{}L_{p} \right)}{\sum_{p \in P}^{}L_{p}}\)
where p is a subperiod of P, Lp is the length of the subperiod p, and Ap is the activity in the subperiod p.
Duration of activities (or proportion of time spent in activities) can be calculated for each activity separately and whatever the time period. If the experimenter decides to group two activities (e.g., lying ruminating and standing ruminating), the duration/proportion of the new activity (here ruminating) is obtained by summing up those of the individual activities.
Number and duration of activity bouts.
Continuous and scan sampling also allow detecting when the activity changes, so that the number of activity bouts can be obtained, and the mean duration of bouts can be calculated. The average bout duration of activity a in bouts collection B is calculated as:
-
\({AvgBoutDuration}_{a}^{B} = \frac{\left( \sum_{b \in B,\ A_{b} = a}^{}D_{b} \right)}{N_{B}(a)}\)
where Db is the duration of the bout b and NB(a) the number of bouts of the collection B where the activity equals a.
Sensor-based systems, however, sometimes provide the time spent in each activity per time period and not the exact timing of a change if any; in this case, the number and the duration of bouts of activity remain unknown.
Calculation of the number of bouts of an activity and of the mean duration of the bouts requires that the recording is done on long periods to avoid edge effects (changes in behaviour that occur at the boundaries of the observed time period). Indeed, when the monitoring starts, the animal is observed in a given activity, but one does not know for how long the animal has been performing the activity. It is common practice to remove the first and the last activity bout observed during the time period studied. The time period must thus be long enough so that several entire bouts of activity can be recorded. In practice, the number of bouts and their mean duration are often calculated per day (see for instance Veissier et al., (2004)).
Assessing the total number of activity bouts in a day, thus, the number of times an animal changes between activities, requires that the ethogram consists of activities described with the same level of details, so that the number of bouts does not depend on which activity an animal performs most during the day. In case of direct observations to answer a specific question, an experimenter may want to sort activities into lying, standing inactive, feeding (all these lasting for minutes or hours) and other activities, that can consist of walking, running, exploring the environment, interacting with other animals or self-directed activities (all of them lasting for a few seconds or minutes). In that case, the number of times an animal changes between activities will largely depend on whether the animal performs the short-lasting activities frequently. The grouping of activities into gross activities should be done before the total number of activity bouts is calculated. Indeed, in the example given above on ruminating while lying or standing, the number of ruminating bouts during a day cannot be calculated by adding the number of lying-ruminating bouts and that of standing-ruminating bouts, because the two activities can be performed in the same bout (i.e., the animal continues to ruminate whilst getting up or lying down).
Duration and number of bouts also largely depend on how bouts are defined. The most common practice is to consider each change of activity as the beginning of a new bout. For example, with continuous observations the number of lying bouts of a cow can be assessed by the number of times the animal lies down; with scan sampling, an eating bout can be defined when eating is observed on at least one scan (Tucker et al., 2009). When scan sampling is used, the interval between scans should be smaller than the duration of activities to not miss bouts. An animal switching from an activity to another can still be considered in the same bout if it returns quickly to the initial activity. One needs to define how long the animal must stop an activity between two instances of that activity so that separate bouts are identified (Yeates et al., 2001).
For instance, eating bouts are usually combined into meals if the interval between successive eating bouts is less than the meal criterion. The minimum interval between bouts can be determined by different methods (Tolkamp et al., 1998, Yeates et al., 2001, Dado & Allen, 1993), for example, using log-survivorship and log-frequency analysis (see Tolkamp et al., (1998), for a description of these methods).
Activity level.
The overall activity in farm animals is usually summarised into an activity level by assigning a weight to each activity, the weight expressing the contribution of the activity to the arousal of the animal (Veissier et al., 2001). The level of activity of the period P is calculated by the sum of the time spent in each activity multiplied by the weight of the activity:
-
\({ActivityLevel}_{a}^{P} = \left( \sum_{\begin{array}{r} p \in P \\ a \in A \end{array}}^{}{T_{ap} \bullet W_{a}} \right)\)
where Tap is the time spent on activity a in period p, Wa is the weight of activity a, the summations are over subperiods p in period P (and over all activities in A).
The weights can be assigned a priori by the experimenter or elicited from observations. Veissier et al. (2001) observed calves to investigate their responses to repeated social regrouping and relocation. In that study, researchers performed a Factorial Analysis of Correspondence (FAC) on the number of instances (scans x calves) of each of five activities per hour; the grouping of factors - that summarises most of the variations between the 24 hours of the day - brought decreasing weights to feeding (1.438), walking (0.763), standing immobile (- 0.085), lying head up (- 0.261), then lying head down (- 0.541), ordering the activities as one would intuitively do to express the decreasing arousal. The FAC is based on associations between activities and therefore, the outcome of the FAC strongly depends on the level of detail of the activities that are included. Experience told us that the ethogram should not be split in too many (short lasting) activities to elicit meaningful weights.
Biological meaning
The time budget of animals depends on their living conditions and the farm management. Cows spend about half of the day lying, but this can vary from 8 h to 13 h (reviewed by Tucker et al., 2021). Lying time is reduced when lying areas are uncomfortable (poorly designed or too hard, wet, small, hot) or not enough resting places are available for the size of the herd (Tucker et al., 2021). Cows may, nevertheless, spend more time lying in case of short cubicles preventing them to stand properly in a cubicle and thus forcing them to lie down as soon as they enter a cubicle (Veissier et al., 2004). The time spent feeding and walking also largely depends on housing and management conditions: cows grazing at pasture spend much more time eating and walking than cows kept indoors and fed herbage harvested from the same pasture (527 min/d eating and 311 min/d walking at pasture vs. 398 min/d and 133 min/d indoor (Dohme-Meier et al., 2014)); bulls and sheep spend less time eating when the diets contain a large proportion of fibre than when the diets contain a large proportion of starch (- 67% in bull fed a 45% starch diet and - 18% in sheep fed a 38% starch diet, compared to animals fed diets with less than 20% starch (Commun et al., 2012; Mialon et al., 2008)). The effects described in the above paragraph are typically observed in all animals from a herd.
Variability is also observed between individuals in a herd. The overall activity of an animal varies over time due to its physiological state. Young cows are often more active and change more often of activity than adult ones (Solano et al., 2016). At the time of oestrus, cows are agitated, spending less time eating but more time active in other ways (more walking, less lying) (Reith & Hoy, 2018). Changes are also observed due to gestation and parturition: the time spent lying by cows decreases during the weeks before and after calving and slowly increases thereafter up to end of lactation, with 2 h of amplitude of variation in multiparous cows (Hut et al., 2022). The changes in activity are generally well marked and short lasting around oestrus but less marked and gradual around calving, making calving detection from gross activity more difficult than that of oestrus (Benaissa et al., 2020).
The overall activity of an animal can also change due to a pathological state (e.g. due to inflammation (Dittrich et al., 2019). These modifications are called sickness behaviour, characterised – among others – by a low activity of the animal (Weary et al., 2009). Metabolic disorders (e.g., hypocalcemia, ketosis, acidosis) are generally accompanied by an increase in the time spent lying and a corresponding decrease in the time spent active and feeding (Weary et al., 2009; Belaid et al. 2021). These changes are more marked in hypocalcemia than in other metabolic diseases, hence the name ‘downer cow’ syndrome for hypocalcemia (Wadhwa & Prasad, 2002). Acidosis can be accompanied by a higher fractioning of activity: sheep suffering from acidosis often change their posture from lying to standing (Commun et al., 2012), as if they do not feel comfortable in either of these postures. Infectious diseases are also associated with an increase in time spent lying down and a decrease in time spent feeding (Weary et al., 2009). Mastitis however may result in a decreased time spent lying down, compensated by an increased time spent standing (Fogsgaard et al., 2015; Medrano-Galarza et al., 2012), presumably due to pain on the udder which is increased by the pressure on it when the animal is lying. Lameness is also accompanied by sickness behaviour and a specific pattern of lying behaviour with less lying bouts, but of longer duration (Solano et al., 2016, de Mol et al., 2013). Sick cows, whatever the origin of the disorders, usually spend less time ruminating (Calamari et al., 2014).
Stress is another factor affecting the activity of animals. Stressed animals usually spend less time lying down and change more often between activities, while distressed animals due to a disease may change between activities less often. For instance, suckling calves separated from their dam and moved from pasture to a barn respond to these changes by spending less time lying down, more time standing still or walking, and by fractionating their activities to a larger extent (Veissier et al., 1989). These modifications fade within days or weeks, indicating habituation to the new environment. Similar changes are observed when primiparous cows join the lactating herd. Around calving, primiparous cows decrease their time spent lying to a larger extent than multiparous cows (Hut et al., 2022) and we suspect that this is due to the many changes undergone by them around calving: they are milked for the first time and they are introduced to the lactating herd and so exposed to social partners and a pen, both novel to them.
Monitoring the overall activity can thus help to check if the animals are managed adequately and to detect physiological states (especially oestrus), pathological states, or stress. Feeding, ruminating and lying seem especially sensitive to variations in the animal or its environment. Lying is also reduced in case of an uncomfortable lying area, so it is often considered that a prolonged time spent lying indicates good welfare (Piñeiro et al., 2019a, 2019b). However, lying time is often increased in case of disease too. In any case, the value obtained for duration and frequency of activities should be interpreted considering the context in which these metrics are obtained, for instance the type and management of feeding, the housing or grazing conditions, and the timing in relation to calving.
Metrics to address fluctuations around the mean activity
Definition
The activity of an animal varies within and between days (Hut et al., 2022). The variation is described by metrics calculating how far values, obtained on a given time frame, are spread around the mean value across several time periods within the time frame. The calculations are generally applied to the duration of activities or the level of activity, less often to the number of bouts or their duration, with all values obtained of individual animals. The time period is often the hour within the day (Mialon et al 2008 (eating duration in bulls), Lardy et al., 2023 (level of activity in cows)) or the day within a period for the actual number of days (Hut et al., 2022 (duration of each activity); Solano et al., 2016 (number of bouts)).
Calculations
The metrics used to describe variations in an animal’s activity across time periods (e.g., hours within a day) are similar to those traditionally used in descriptive statistics except that they are applied at individual level (to characterise the variability of the activity of a given animal and not the variability between animals):
Minimum (Min) refers to the minimum value observed/recorded
Maximum (Max) refers to the maximum value observed/recorded
Range corresponds to the difference between Min and Max
Quantiles
Quantiles are cut points dividing the dataset into continuous intervals with equal probabilities. The most commonly used quantiles are quartiles, which divide the number of data points into four parts, where the first quartile (Q1, 25th percentile) is the maximum value in the 25% of the lowest values in the dataset, the second quartile (Q2) corresponds to the median, and the third quartile (Q3, 75th percentile) is the minimum value in the 25% of the highest values in the dataset.
Variance and Standard deviation
The variance is the sum of the squares of the differences between each value and the mean (see formula below); and Standard deviation (SD) is the square root of the variance.
-
\(Variance(x) = \ \frac{1}{N - 1}\sum_{i = 1}^{N}\left( x_{i} - \mu \right)^{2}\)
where x is the activity (expressed for examples as the level of activity or the proportion of time spent in a given activity) composed of N observations, xi is the ith observation, and µ the mean of the activity.
Root Mean Square of the Successive Differences (RMSSD):
RMSSD measures the variations from one interval to the next one. RMSSD is calculated as follows:
-
\(RMSSD(x) = \ \sqrt[2]{\frac{1}{N - 1}\sum_{i = 1}^{N - 1}\left( x_{i} - x_{i + 1} \right)^{2}}\)
where x is the sequence of activities (expressed for example as the level of activity) composed of N observations, xi is the ith observation.
Irregularity
An index of irregularity of behaviour can be calculated as proposed for feed intake (Salgado et al., 2021a, 2021b). First, a regression line of cumulative data (e.g., cumulative time spent in a given activity) is drawn, then the differences between observed values and the regression line are calculated. The sum of these differences brings the irregularity index.
Biological meaning
Maximum activity during the day can be relevant to identify cow states: in a study where many metrics were used for a random forest classification (i.e., multiple decision trees created using different random subsets of the data), Lardy et al. (2023) found that the maximum activity during the day and Quantile 90 (two metrics closely linked) were the most important features to discriminate pathological and physiological states of cows.
Within-day variations of activity are well marked in healthy and non-stressed animals. For instance, lying is predominant at some time points throughout the day whereas eating and other activities are predominant at other times. Therefore, the activity level is not constant from hour to hour during the day. Within-day variations (measured by SD and RMSSD) are less marked in sick animals than in healthy ones; this is the case for cows affected by mastitis and to a lesser extent by lameness (Veissier et al., 2017). Within-day variations are also affected by oestrus: variations rise above baseline at the beginning of oestrus then decrease below baseline for at least two days (Veissier et al., 2017). RMSSD slightly differs from SD. With the same amount of variation during the day (same SD), an activity that varies smoothly between successive hours results in a low RMSSD while an activity that fluctuates between successive hours results in a high RMSSD. In Veissier et al. (2017) the decrease in the within-day variation in the activity of cows affected by mastitis, lameness and oestrus was more marked when assessed by RMSSD than by SD, suggesting that cows change of activity more often when diseased or in oestrus. Stress may also be associated with a reduction in within-day variation in activity, as observed in cattle moved from pasture to indoor housing (Veissier et al., 1989; using the difference between night and early morning). Spreading activities throughout the day can also be a way to adapt to specific conditions. Bulls fed fibrous diets eat in few meals during the day whereas bulls fed high starch diets spread their eating activity over the entire day, which results in a low SD of eating duration (Mialon et al., 2008). Dispersing small meals over the day is likely to be a strategy to avoid ruminal acidosis due to high amounts of starch in a diet.
Between-day variation in activity can increase when animals are disturbed as a result of disease or in case their activity pattern is interrupted by sudden events. For instance, cattle and sheep affected by acidosis or ketosis have more variable activities across days than healthy animals (Commun et al., 2012; González et al., 2008). At least in case of ketosis, an effective treatment eliminates the effect (Goldhawk et al., 2009). Lame cows can also display high between-days variation in the duration of lying bouts (Ito et al., 2010; Solano et al., 2016). A quick return to normal or baseline values after small disturbances (micro-recoveries) results in low variance, and is considered as a sign of good resilience (Scheffer et al., 2018). Animals that spontaneously (i.e., apart from diseases or other challenges) have a variable activity are less prone to further diseases (Van Dixhoorn et al., 2018).
Metrics to address the cyclicity of activity
Definition
Cyclicity indicates fluctuations at regular intervals around activity trends. The most common cycle is the circadian cycle, which is the 24-h rhythm due to the alternance of day and night.
Calculations
Several metrics can be used for addressing the cyclicity of activity (Kok et al., 2023). Autocorrelation and Fourier transform are basic calculations. Other indices are derived from these two metrics such as non-periodicity index (Van Dixhoorn et al., 2023; Van Dixhoorn et al., 2018) and degree of functional coupling (Berger et al., 2003; Scheibe et al., 1999).
Autocorrelation
Autocorrelation measures the correlation between successive values of a signal. The autocorrelation depends on the chosen delay, e.g., for hourly data the lag-1 corresponds to the correlation between successive hours and lag-24 corresponds to the correlation between an hour of the day and the same hour of the next day, therefore reflecting the circadian cycle. Let x be the number of measured behaviour events (e.g. duration, frequency, etc), and “ l “ the amount of time that has passed (lag l). For a sequence x of size N, the autocorrelation function (ACF) with a lag l is calculated as following:
-
\(ACF(x,l) = \ \frac{1}{(N - l)\sigma^{2}}\sum_{i = 1}^{N - l}{\left( x_{i} - \mu \right)\left( x_{i + l} - \mu \right)}\)
Where σ is the variance of the sequence x, μ is the average value of the sequence x and xi the ith element of the sequence x.
Non-periodicity
Non-periodicity is calculated by plotting the correlogram of the raw data (which is a graphical display of a correlation matrix of the data) over a sinusoid with an amplitude of 0.25 and a 24-hour cycle and assessing the difference between the correlogram and the sinusoid by calculating the mean squared error (Figure 2) (Van Dixhoorn et al., 2023; Van Dixhoorn et al., 2018). The value of an amplitude of 0.25 is chosen as it gave the best fit and might be adjusted in other situations where the autocorrelation shows a circadian rhythm.
-
\(Nonperiodicity(x) = \frac{\left( \sum_{l = 1\ldots 100}^{}\left( ACF(x,l) - 0.25 \bullet \cos\left( \frac{2\pi \bullet l}{24} \right) \right)^{2} \right)}{100}\)
where x is a variable measured at hourly (or other chosen time interval) time intervals, ACF(x,l) is the autocorrelation function for variable x at lag l (ranging from 1 to 100), 0.25∙cos(2π∙l/24) is the cosine function with a 24-h cycle and an amplitude of 0.25 that is used as a fit function.
Fourier transform
Fourier transform represents the sinusoids that compose the original variation. Each sinusoid is defined by a frequency and an amplitude. The contribution of each sinusoid to explain the original variation is expressed in absolute or relative power, usually referring to the frequency of the sinusoid or to frequency bands. For instance, the contribution of the circadian cycle and of ultradian cycles can be calculated. When variations within 24 h time series are analysed by Fourier transform, the fundamental (h0) refers to the average activity during 24 h, harmonic 1 (h1) refers to variations following a 24 h cycle (once per day); h2, to a 12 h cycle (twice per day); h3, to an 8 h cycle (3 times per day); h4 to a 6 h cycle (4 times per day), etc. The main cycle is the circadian one; the activity of an animal can therefore be modelled by its overall activity (mean during 24 h) and the variations around overall activity following a 24 h cycle, in other words into h0 and h1. The Fourier-based approximation with thresholding (FBAT) method was developed to compare such models obtained on successive time series (Wagner et al., 2021). An alternative to the Fourier transform is the Cosinor method. Cosinor and Fourier are analogous in formulation, but differ in operation (see (Chkeir et al., 2019), for a comparison of the two modelling approaches). An example of Fourier transform is visualised in Figure 2.
Degree of functional coupling
The degree of functional coupling (DFC) is obtained by calculating autocorrelations, then applying Fourier transform to the correlogram, extracting the significant harmonics and calculating the power of each significant harmonics out of the power of all significant harmonics. More specifically, the relative power of the harmonic corresponding to a 24 h cycle expresses how much the variations are due to the circadian cycle: when DFC equals 100%, the variation in activity follows strictly a circadian cycle vs. when DFC equals 0%, the activity does not at all depend on the 24 h cycle (Berger et al., 2003).
In theory, data during 24 h only can be used to identify a circadian cycle. In practice, activity data usually contains noise, i.e., erratic fluctuations so that more than one day is necessary to identify correctly cyclic components. The number of days required depends on the amount of noise vs. cyclic components.
In general, the activity of an animal peaks several times during the day, e.g., depending on the timing of feeding or milking, whereas the activity stays low at night. Autocorrelations and Fourier transform hardly take the dissymmetry between day and night activity into account. Wavelet methods could be used to overcome this problem (discussed in Wagner et al., 2021).
Biological meaning
Daily periodicities are influenced by internal clocks (endogenous driven biological cycles) and external factors such as temperature, light, humidity, feeding time (exogenously driven biological cycles) (Saper et al., 2005). Internal clocks generate a rhythm of about 24 h period. In mammals, the central internal clock situated in the brain (suprachiasmatic nuclei) coordinates peripheral clocks in the body (Honma, 2018). Among external cues (or “Zeitgebers”), light is known to be the most powerful one, impacting both on behaviour and physiological functions (Honma, 2018). For instance, cows are typically diurnal animals: they eat essentially between dawn and dusk and they predominantly rest at night (DeVries et al., 2003; Hafez et al., 1969). Variations can nevertheless be observed between cows in the cyclicity of their behaviour. Competition for resources (e.g. feed, lying area), due to overstocking or ambiguity in the ranking order because of frequent change in group composition, can cause a misalignment with the circadian rhythm (McCabe et al., 2021; Van Dixhoorn et al., 2023; Van Dixhoorn et al., 2018; Van Erp et al., 2020).
Cows with more marked circadian patterns of activity seem more resistant to health disorders. For instance, cows with marked cyclicity of eating, walking or lying before calving are less affected by post-partum health disorders (including inflammatory and metabolic problems (Van Dixhoorn et al., 2023; Van Dixhoorn et al., 2018)). Indeed, dairy cows need to have their physiological mechanisms fine-tuned to be able to produce large quantities of milk while avoiding nutritional and metabolic deficiencies (negative energy balance or mineral deficiencies such as hypocalcemia). We hypothesise that good cyclicity, aligned with circadian rhythm can help to avoid such dysfunctions, especially in the demanding postpartum period.
Any change of the internal state of an animal - due to stress, disease or some specific reproductive status (parturition, oestrus) – can in turn affect the cyclicity of activities. For instance, the difference between activity during the day and at night is less marked in heifers experiencing a large change in their environment – weaning and turning from pasture to indoors – and in cows affected by mastitis or lameness (Veissier et al., 1989; Veissier et al., 2017). Variations during the day can be modelled, e.g., thanks to Fourier transform in the FBAT method. The distance between models obtained on successive 24 h series increases when cows are stressed, diseased, in oestrus or about to calve, expressing a change in daily patterns (Wagner et al., 2021). These effects may be due to the release of glucocorticoids during stress, disease or even calving. Indeed, glucocorticoids, the secretion of which follows a circadian pattern, help to coordinate peripheral clocks with the brain pacemaker (Dumbell et al., 2016).
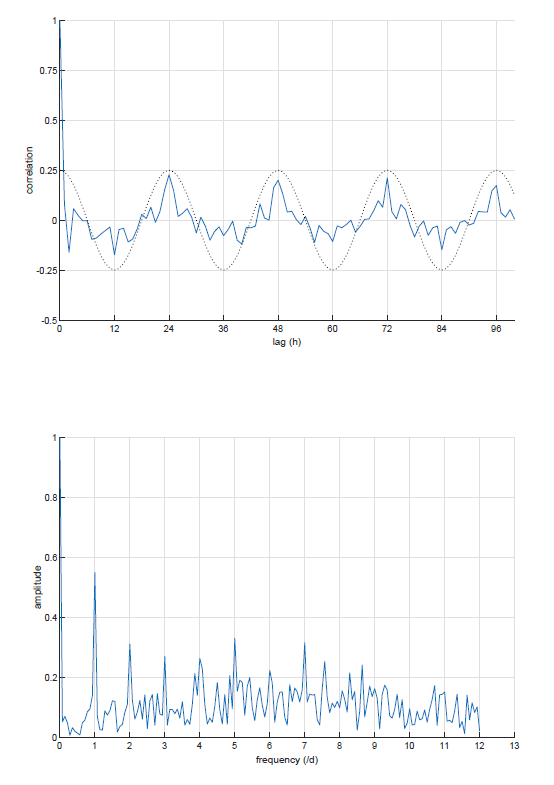
Figure 2 - Examples of non-periodicity of eating behaviour visualised through correlograms (top), and Fourier transform pattern (bottom) (frequency of activity performed per day). In the correlograms, dotted lines represent the expected cyclicity of the specific behaviour, while the solid lines indicate the observed behaviours. Non-periodicity is assessed by calculating the Root Mean Squared Error (RMSE) of the correlogram as compared to the sinusoid. In the Fourier transform pattern the amplitude is given per frequency, expressing the strengh of the cycles in activity for that frequency. The peak at frequency 1 shows that this cow has a strong circadian pattern. The sum of the amplitudes at frequency 1, 2, 3 and 4 represents the strength of the cycles in activitities with a 24, 12, 8, and 6 h cycles and is used as a measure of the cyclicity of the cow.
Metrics to address the synchrony between animals
Definition
Synchrony measures the extent to which animals of a given group perform the same activity at the same time. The synchrony can be assessed between two animals, between an animal and the group it belongs to, or at the group level.
Calculations
Synchrony is to be assessed based on what animals do at certain points in time spaced at regular intervals. Collecting data using scan sampling is therefore appropriate for calculations of synchrony metrics. In case of continuous observations, where changes in behaviour are noted for each animal exactly when they occur, a pre-processing of data is necessary: at each time interval the instantaneous activity of each animal of the group need to be extracted (resampling procedure). When data are expressed as main activity of each animal during a certain interval, as often done with sensors, synchrony can only be approximated: if the intervals at which the activity is noted are short enough (e.g. 5 min) then it may be considered that the animal kept the same activity during the whole interval, and the data can then be processed as scan sampling.
Several metrics can be found in the literature to calculate synchrony. We cite here the most common ones.
-
The synchrony between two animals is calculated as the proportion of the scans during which they are engaged in the same activity (see example in Veissier et al., 1989):
-
\(synchrony = \ \frac{1}{n}\sum_{i = 1}^{n}S_{i}\)
where n is the number of scans, Si equals 1 if the two animals have the same activity at scan i and 0 if they do not.
The synchrony of a focal animal (i.e., the animal observed) with the rest of the group or a certain sub-group of animals in the group can be calculated as:
-
the average of the proportion of animals from the group or the sub-group performing the same activity as the focal animal across scans. This also corresponds to the average of synchrony indices (a) obtained for a focal animal and any other animal from the herd or the sub-group (Veissier et al., 1989):
-
\(synchrony = \ \frac{1}{n(m - 1)}\sum_{j = 1}^{m - 1}{\sum_{i = 1}^{n}S_{ij}}\)
where n is the number of scans, m is the number of animals in the group, Sij equals 1 if the focal animal and the other animal j have the same activity at scan i and 0 if they do not.
-
the proportion of scans during which the focal individual performs the same activity as most individuals of the rest of the group (Ruckstuhl, 1999). The calculation is similar to that for the synchrony between two animals given in (a) above but with Si equals 1 when the focal animal has the same activity as most of the group and 0 when it does not.
At group level, the synchrony can be calculated with several indices:
-
The proportion of scans where all animals of the group perform the same activity; Again, similar calculations as for (a) are used with Si equals 1 if all animals perform the same activity and 0 if not.
-
The average of metrics (b) (Veissier et al., 1989) or (c) (Asher & Collins, 2012) that represent the average synchrony calculated across all animals in a group. If all animals perform always the same activity, the result will be 1 with metrics (d) and (e). If not, metrics (e) will provide higher and values than metric (d). for instance, if animals never perform all the same activity, metric (d) will be 0 whereas metrics (e) can still give an idea of how many animals perform the same activity.
The metrics presented above depend largely on the number of activity categories and the number of animals in the group (especially metrics (d)): when the group is large and the number of activity categories is high, there is little chance that animals perform the same activity at the same time. Asher and Collins (2012) thus recommend comparing the distribution of activity observed with the one obtained at random. This can be done with:
-
Kappa coefficient of agreement. For instance, the proportion of pairs of animals observed with the same activity is calculated (Rook & Penning, 1991):
-
\(P(O) = \ \frac{1}{np}\sum_{i = 1}^{n}S_{i}\)
Where capital P refers to proportion, n is the number of scans, p is the total number of pairs of animals in the groups, and Si the number of pairs of animals performing the same activity at scan i.
The expected proportion of pairs that would perform the same activity by chance is then calculated:
-
\(P(E) = \ \frac{1}{(nm)^{2}}\sum_{k = 1}^{l}C_{k}^{2}\)
where n is the number of scans, m is the group size, l is the number of activity categories, and Ck the frequency of observation of activity category k (i.e., total number of scan x animals occurrence of the activity).
Then
-
\(Kappa\ coefficient = \ \frac{P(O) - P(E)}{1 - P(E)}\)
The Kappa coefficient equals 1 if all animals always perform the same activity at the same time (full synchronization). It equals 0 when animals are not synchronised more than at random.
-
Other methods can be found in the literature to compare the synchrony observed to that expected on a randomised dataset: calculating a dispersion index (Raussi et al., 2011) or applying Monte-Carlo methods (Whitehead, 1999). These two options are rarely used in the literature on behaviour (e.g., we did not find studies using the dispersion index apart from that of Raussi et al. (2011)). In most cases, these methods may not have added value compared to the Kappa coefficient of agreement.
The activity categories should be carefully chosen. If there are too many categories (e.g., detailing precisely what the animals do when standing active: walking, scratching, interacting with each other), the animals will seem little synchronised whereas if there are too few categories (e.g., active vs. inactive) they will seem very synchronised. Asher and Collins (2012) recommend using 5 activity categories in laying hens. It must be considered whether or not we expect social facilitation of an activity to occur (that is one animal engaging in an activity leads to other animals engaging in the same activity). In ruminants, ruminating appears as a reflex activity, ruminating thus should not be used as a separate activity but rather included in lying and standing idling (i.e., postures when ruminating can occur) because we do not expect social facilitation of ruminating.
To be interpreted in terms of a positive relationship between two animals, the synchrony needs to be estimated between animals that have about the same time budget (i.e., same amount of time spent in each activity per day). For instance, although a cow has a strong bond to its new-born calf, the apparent synchrony between them may be low because the cow spends lot of the time foraging whereas the calf spends more time lying (Veissier et al., 1990).
Biological meaning
Animals may be synchronised because the activity of an animal is influenced by that of other animals. Social facilitation has been described in many species and contexts (Clayton, 1978). Animals may be synchronised also because they adopt a similar rhythm of activity: activities follow a circadian rhythm triggered at least in part by external cues such as light or timing of food distribution (or milking in dairy cows) so if animals are subjected to the same cues, their activity will tend to be similar (Flury & Gygax, 2016).
The synchrony between two animals (fighting excluded) gives us an estimate of how closely (and positively) they are related to each other. Two animals bond by a positive social relationship have more chances than unrelated animals to express the same activity at the same time. For instance, when calves stay with their dam after weaning they keep preferential relations that are shown by proximity, exchanges of positive interactions, and also synchrony (Veissier et al., 1990).
Synchrony at the group level reflects social cohesion, i.e. the strength of the bonds between the animals in the group (Clayton, 1978). Groups newly formed are usually less cohesive, with animals exchanging aggressive interactions and being less synchronised (Mounier et al., 2005). The synchrony can also decrease if there is competition for access to a resource, e.g., food and lying places. For instance, in cows and sheep, synchrony of lying decreases when lying space is limited (Bøe et al., 2006; Winckler et al., 2015). The synchrony between an animal and the rest of the group reflects the familiarity of that animal with the group: synchrony may be low in case of a newly introduced animals until the organization of the group is stabilised (Arey, 1999; Boyle et al., 2013). A variation in synchrony can also be caused by a health disorder: the activity of an animal is modified in case of disease (Dantzer et al., 2008), so that the diseased animal can depart from the rest of the group (e.g., a cow isolates and stay standing idling or lying for longer when ill (Proudfoot & Habing, 2015; Proudfoot et al., 2012).
Discussion
In this paper we present metrics that can be calculated from data on gross activity and identify their conditions for use (summarised in Table 1). We divided the metrics into four groups that represent different aspects of animal behaviour: the overall activity, the fluctuations around the mean activity, the cyclicity, and the synchrony between animals.
The overall activity is the most often studied aspect. It represents the total duration and organisation of the activity in bouts. The fluctuations of activity during the day or across days and the organisation of these fluctuations according to (circadian) cycles are less often addressed but are gaining attention in research. Synchrony between animals is generally used to study the social organisation of a group of animals, but is also more and more used as specific indicators (e.g. likelihood of diseases or evaluation of the management) and constitutes also a promising indicator of positive welfare (Keeling et al., 2021; Napolitano et al., 2009).
These four groups of metrics can be used to analyse the impact of housing and management procedures. In that case, the values at herd level are evaluated, all animals within the herd are taken into account. In most cases, the average herd level values per metric are expected to not vary much in time, unless management or housing change. However, seasonality may affect some metrics due to photoperiod, weather, or the reproductive period (e.g. females may be less synchronised with the rest of the group when they have young).
These metrics can also be used to compare animals within a herd individually (with same management and housing). Animals are usually consistent with time, so that the data can be used to phenotype them (Bacher et al., 2022; Poppe et al., 2022). In turn the behavioural phenotype can inform about other traits. For instance, a cow that shows low regularity in activity is likely to be more susceptible to post-partum diseases, when she has to cope with metabolic constraints (van Dixhoorn et al., 2023).
Transient changes in activity can be observed under certain circumstances at animal level, especially when an animal is sick, in a specific physiological state (oestrus, calving), or stressed. When such transient changes are observed concurrently in most animals of the herd, it is likely that the herd has been disturbed by external events (e.g., handling to apply a treatment, hoof trimming, period of heat stress). When the transient change is observed in only one or few animals, it is more likely an individual case of disease or a reproductive event.
Table 1 - Summary of metrics to describe the activity of animals with their condition of use
|
Category |
Purpose |
Metrics |
Raw data |
Limitations |
Reference |
|---|---|---|---|---|---|
|
Overall activity |
Detection of physiological states (e.g. oestrus), pathological states (e.g. lameness), or stress |
Proportion of time spent in an activity |
Collection of activity bouts with duration |
Depends on their living conditions and the farm management. Modifications are also observed between individuals. There is a need to :
|
|
|
Number and duration of activity bouts |
Collection of activity bouts with duration |
||||
|
Activity level |
Time spent on activities |
Weights of each activity to be elicited |
|||
|
Fluctuations around the mean |
To identify pathological or physiological statesc |
Min/max/range/quantiles/variance/RMSSD |
Scan sampling and continuous sampling |
||
|
Irregularity |
Continuous sampling |
||||
|
Cyclicity |
Detection of physiological states (e.g., oestrus), pathological states (lameness), or stress. To predict resilience. |
Autocorrelation |
Continuous sampling |
Recording for long periods (longer than the cycle to be detected) |
van Dixhoorn et al., 2023, 2018 |
|
Non-periodicity |
Continuous sampling |
van Dixhoorn et al., 2023, 2018 |
|||
|
Cosinor method |
Continuous sampling |
||||
|
Fourier transform |
Continuous sampling |
van Dixhoorn et al., 2023 |
|||
|
Degree of functional coupling (DFC) |
Continuous sampling |
||||
|
Synchrony between individuals |
To detect bonds between animals and / or availability of resources. Inclusion of an animal in a group. Detection of health disorders |
Proportion of animals from the group or sub-group performing the same activity as a focal individual |
Scan sampling |
Depends on the number of animals and number of activity categories |
Veissier and le Neindre, 1989 |
|
Proportion of scans during which a focal individual performs the same activity as most individuals of the rest of the group |
Scan sampling |
Ruckstuhl, 1999; Asher and Collins 2012 |
|||
|
Synchrony at herd level |
Social cohesion, competition for resources |
The average of metrics taken at individual level |
Scan sampling and continuous sampling |
Depends on the number of animals and number of activity categories |
Veissier et al., 1989; Stoye et al., 2012; Arsher and Collins, 2012 |
|
The proportion of scans where all animals of the group perform the same activity |
Scan sampling and continuous sampling |
Stoye et al., 2012; Arsher and Collins, 2012 |
|||
|
Kappa coefficient of agreement |
Scan sampling and continuous sampling |
Rook and Penning, 1991 |
Combining several behavioural metrics is usually necessary to have a comprehensive overview of the internal state of an animal. For instance, to infer the internal state of a cow (diseased, in oestrus, about to calve, or stressed), metrics on overall activity, fluctuations and cyclicity are necessary to be able to classify the cow into the corresponding state (Lardy et al., 2023).
In this paper, only metrics describing overall activity, fluctuations around mean activity, cyclicity, and synchrony between animals are described and discussed. The data can nevertheless be further processed to extract more information. For instance, a network analysis could be performed with links between individuals of a group estimated from their synchrony. Such an approach could probably be applied to study group effects such as social facilitation, leadership, or cooperation among individuals. Markov chains can be used to analyse sequences of activities to better understand the organisation of activities (Rugg & Buech, 1990; Schafer et al., 2020). Machine learning applied to metrics describing activity can also help to classify animals according to their phenotype or to detect changes in activity for specific animals and days (Lardy et al., 2023; Wagner et al., 2020, Debauche et al., 2021).
In conclusion, activity data provide the raw material for the calculation of several metrics that describe animal behaviour. The choice of which metrics to use, depends on the research question or potential application. A clear research question is essential for the selection of the most appropriate metrics that best characterise specific aspects of the behaviour of the animals, suitable for answering question(s) asked. We believe that clarification of the metrics and on how they should be calculated will help to standardise these metrics, making them easier to use and allowing comparisons between studies.
Acknowledgments
Preprint version 5 of this article has been peer-reviewed and recommended by Peer Community In Animal Science (https://doi.org/10.24072/pci.animsci.100268; Chincarini, 2024).
Funding
The study received financial support via the SmartCow project funded by the European Commission (Horizon 2020 research and innovation program, grant agreement No. 730924) and is part of IRC-SAE (CAP 20–25).
Data, scripts, code, and supplementary information availability
Examples of calculations of metrics using R software are available online (https://doi.org/10.6084/m9.figshare.24891252; van Dixhoorn, 2023).
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article. Isabelle Veissier is recommender of PCI Animal Science.