
 CC-BY 4.0
CC-BY 4.0
Introduction
Benthic landscape homogenization reduces community resilience (Cimon and Cusson, 2018), and has therefore become a research priority for marine biodiversity conservation. Seagrass meadows provide temporal, spatial, structural and functional complexity, and are among the most productive coastal ecosystems (Unsworth et al. 2022). Although they cover less than 0.2% of the ocean’s surface, seagrass meadows play thus an important ecological role, providing valuable services to coastal ecosystems and people (Fourqurean et al., 2012; Unsworth et al., 2022). As ecosystem engineers (Bos et al., 2007; van der Heide et al., 2012), seagrasses provide a stable habitat for many species (Hughes et al., 2009), they attenuate hydrodynamic energy from waves (Paul and Amos, 2011) and currents (Widdows et al., 2008), and sequester carbon (Fourqurean et al., 2012; Macreadie et al., 2021). These meadows however are one of the most threatened marine habitats, with a global decline of 29% since 1980 due to rapid coastal development and associated problems such as eutrophication (Dunic et al., 2021; Turschwell et al., 2021).
In the northern hemisphere, seagrass meadows are dominated by two species, Zostera marina Linnaeus, 1753, and Zostera noltei Hornemann, 1832, and extend from the northern Atlantic to the coast of Mauritania (https://www.gbif.org, Short et al., 2007). Unlike algae, these flowering plants with stems and long green leaves have roots and rhizomes that anchor them to sandy or muddy seabeds and allow them to extract nutrients from both the water and the sediment. The two species differ in their ecology, spatial distribution and responses to environmental changes (Moore and Short, 2006; Ouisse et al., 2012, 2011). Thanks to its high tolerance to desiccation and high temperature, beds of Z. noltei can be found in the intertidal zone or in the shallow subtidal zone of lagoon areas (Borum and Greve, 2004). Z. marina forms predominantly subtidal meadows, down to 40 m depth in the clearest waters but reaching only a few meters in the most turbid areas (Duarte, 1991). The population dynamics of these two species are largely controlled by seasonal variations in temperature, light and nutrient (Lee et al., 2007). Z. noltei is a perennial plant with limited leaf cover during winter (typically one leaf per shoot), leaving sometimes only the below-ground biomass (Larkum et al., 2006). In low/southern latitudes, some populations may continue to grow under mild winter conditions with a slight decrease in light and temperature. Z. marina is a perennial plant that tends to exhibit optimal productivity and growth rate during the warmer months with longer days (Lee et al., 2007).
Both species are characterized by a high morphological plasticity. Within species, population plasticity reflects processes of acclimatisation to large-scale and/or local environmental conditions, which influence their resilience and ability to survive. Changes are observed at different levels, from individual physiology to population dynamics. For example, low shoot density and higher aboveground biomass relative to roots and rhizomes at low light levels are known responses of seagrasses to reduce self-shading (Krause-Jensen et al., 2000; Short et al., 1995), thereby promoting photosynthetic production over respiration (Olesen and Sand-Jensen, 1993). As another example, seagrasses can adjust their tissue nitrogen or carbon levels and shoot density under nutrient stress (Roca et al., 2016). Despite these adaptive capacities, severe losses of European meadows have been documented in recent decades in response to human-induced or natural environmental degradation (Ouisse et al., 2020; Plus et al., 2010; Unsworth et al., 2017), although encouraging trends have been observed in some areas (de los Santos et al., 2019; Dunic et al., 2021). Ongoing global change suggests an increased vulnerability of these habitat-forming species in the future, which may increase the risk of local extinctions (Turschwell et al., 2021). In this context, it is essential to further describe and understand the ecology of seagrass meadow species and to characterise their trajectories across their spatial range in relation to environmental constraints.
In order to monitor, detect, quantify and predict temporal changes in biological communities, methods are needed that can link temporal patterns to ecological processes. Community Trajectory Analysis (CTA ; De Cáceres et al., 2019) is a recent multivariate method that can describe single trajectories by quantifying the changes between successive observations, the direction of these changes or the overall dynamics of the community. In CTA, closely spaced successive observations would suggest the presence of stable communities characterised by non-directional and gradual changes. Directional changes (i.e. trajectory following a particular direction) or saltatory changes (sudden and abrupt change between successive observations) would imply succession or regime shifts following a disturbance (Matthews et al., 2013; Toumi et al., 2023). In addition, this tool allows trajectories to be compared and the spatial variability of community dynamics to be captured.
Using the CTA framework, this study aimed to assess how the population and morphometric traits of two iconic seagrass species (Z. marina and Z. noltei) vary seasonally at a regional scale, in relation to environmental conditions. To date, most published studies covering large environmental gradients have limited temporal replication (e.g. Soissons et al., 2018a; Yang et al., 2013). We also investigated fine-scale variability of the habitat (i.e. within meadows), considering that seagrass meadows are not homogeneous structures (Brun et al., 2003), but host individuals that continuously colonize and vacate patches to the rhythm of morphometric development, recruitment and mortality, which scale up to habitat expansions or contractions. To our knowledge, this is one of the few studies to propose such a spatial and temporal scale of simultaneous observations for these two species. The results will increase our ecological knowledge of Zostera spp., which is essential to guide management decisions and improve conservation efforts.
Materials and methods
Site description
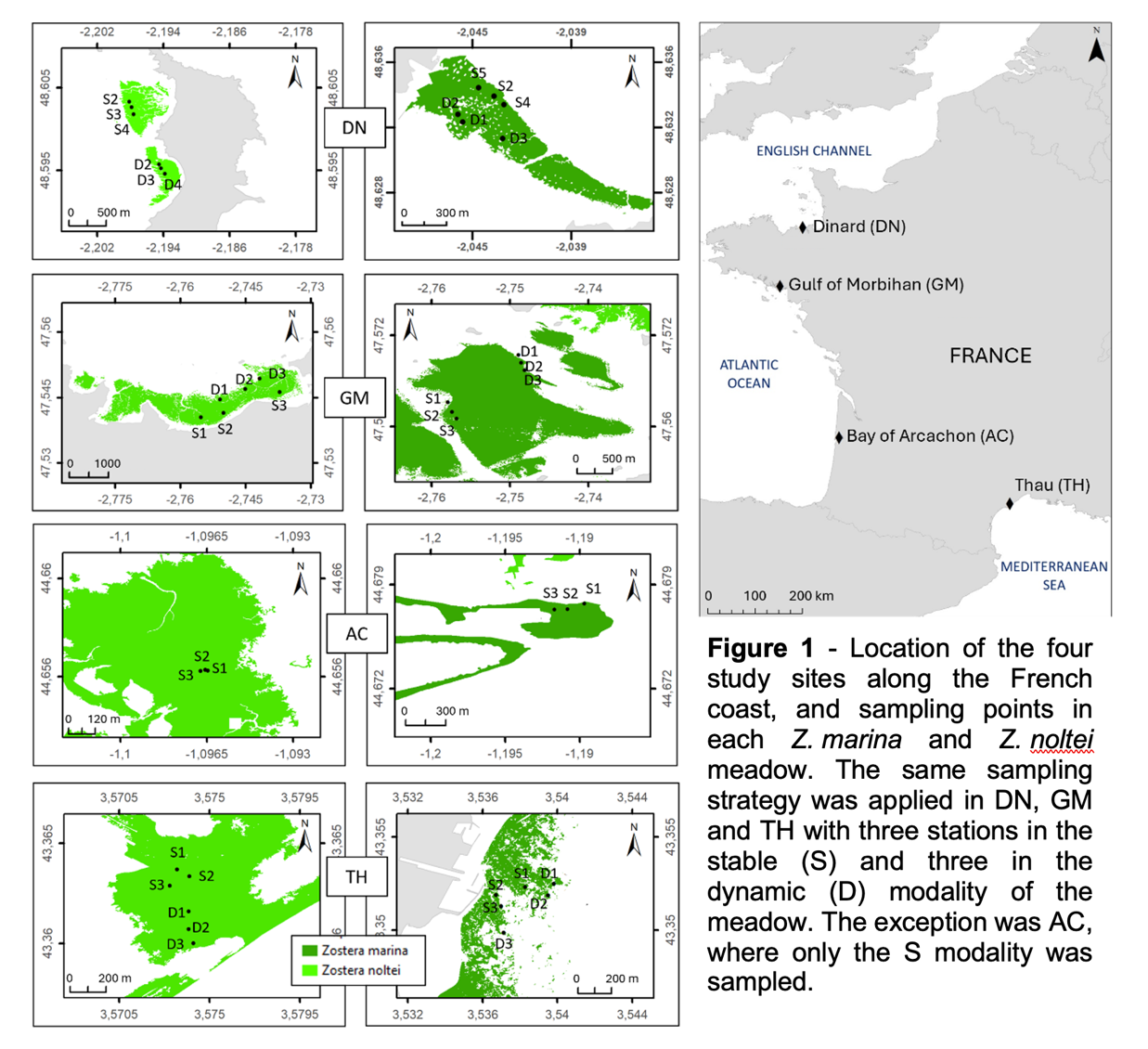
Zostera beds were monitored in four contrasting coastal sites in the English Channel, the Bay of Biscay and the Western Mediterranean (Fig. 1). Thau (hereafter TH), the Mediterranean southernmost site, differs from the others in that it is a semi-enclosed lagoon (68 km2) with a microtidal regime. This site exhibits the widest water temperature range compared with other sites, with the highest values in summer and the lowest values in winter (Table 1). This site is also the one with the highest salinity and lowest mean turbidity (Table 1) and hosts exclusively subtidal seagrass meadows (both Z. marina and Z. noltei). The shallow semi-enclosed bay of Arcachon (hereafter AC) in the Bay of Biscay has a total area of 174 km2 with 70% of intertidal flats. These mudflats are colonized by Z. noltei while Z. marina occupies the shallow subtidal sector around the channel edges. This site is the most eutrophic of our four study sites, with the highest mean annual chlorophyll-a (chl-a) and ammonium (NH4+) levels. It also has the most turbid waters (Table 1). Further north in the Bay of Biscay, the semi-enclosed Gulf of Morbihan (hereafter GM) occupies 115 km2 separated from the sea by a 900 meter wide inlet. The sampled meadows were mostly located in the eastern part of the gulf, in a sheltered area. At this site the chl-a concentration is low during most of the year, with a spring bloom (Table 1). Turbidity may reach very high values in winter (Table 1). The northernmost site of Dinard (hereafter DN) lies in the English Channel. At this site, the Z. noltei bed is located in the intertidal zone of a semi-enclosed bay to the west of DN. The studied Z. marina bed is located at the mouth of the Rance river, exposed to a continuous and intense tidal flow with periods of emergence during the spring tides. This site has the lowest mean annual temperature, and intermediate salinity compared with the other sites. Annual levels of NH4+ are low, as is chl-a, except during the spring bloom (Table 1).
During the course of the study, water temperature (waterTemp) and light were measured continuously using data loggers (Onset HOBO Pendant Temperature/Light Data Logger), in each habitat (each site, Z. marina and Z. noltei separately). To correlate these environmental variables with seagrass traits, data were averaged over two months preceding the sampling date in order to match the time-scale of seagrass dynamics (Roca et al., 2016). Light intensity was averaged on a daily basis, considering only the daytime period of immersion (waterLight) or both immersion and emersion (light total for Z. noltei). Salinity, nutrients (NH4+: ammonium, PO43-: phosphate) and chl-a in water were measured at the bed scale (Z. marina and Z. noltei) at each site and each season. Sediment cores (5 cm2) were collected for sediment organic matter content estimation (OM) by loss on ignition method (520°C for 6h). Details on these methods are available in Lacoste et al. (2024).
Field sampling
At each site and for each species (Z. marina and Z. noltei), two locations were sampled (hereafter referred to as Modalities; Fig. 1). They were defined based on historical maps and/or aerial photography when available, or on local expert knowledge. The first modality aims at representing the most stable area of the meadow (S), namely an area occupied by seagrass over, at least the last decade. The second modality targets a dynamic part of the meadow (D), i.e. an area subject to fluctuating seagrass colonization over time, which is presumed to be more unstable. In AC, no distinction could be made between the two modalities due to the specific configuration of the seagrass beds which have been in sharp decline since 2000 (Plus et al., 2010). The sampled area was considered to be the stable modality where seagrass persists. Each site was visited seasonally between winter 2020 and spring 2021 (see details in Lacoste et al. 2024), except in spring 2020 due to the Covid-19 pandemic. Other exceptions were in winter 2020 in GM and TH for Z. marina, due to weather conditions limiting site accessibility. For each Site × Species × Season × Modality combination, samples were collected in three stations (see next section).
Table 1 - Seasonal variations (mean ± sd) of sub-surface salinity, water temperature, turbidity, ammonium (NH4+) and chlorophyll-a (chl-a) concentrations at the four study sites with the seagrass bed surface area and tidal regime. (DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau ; ZM: Zostera marina, ZN: Zostera noltei). Data were extracted from the monitoring program REPHY (2022, https://www.seanoe.org/data/00361/47248) and averaged seasonally over six years (2016-2021).
SITE | Season | Salinity (PSU) | Temperature (°C) | Turbidity (FNU) | NH4+ (µmol.l-1) | chl-a (µg.l-1) | Bed size (ha)* | Tidal regime | |
|---|---|---|---|---|---|---|---|---|---|
ZM | ZN | ||||||||
DN | Winter | 34.8±0.2 | 10.0 ± 0.9 | 3.7 ± 0.9 | 0.27 ± 0.24 | 0.72 ± 0.34 | 217 | 27 | mega |
Spring | 34.8 ± 0.1 | 11.0 ± 1.9 | 2.4 ± 2.1 | 0.12 ± 0.08 | 2.75 ± 1.76 | ||||
Summer | 34.9±0.1 | 17.4 ± 1.8 | 0.7 ± 0.1 | 0.07 ± 0.04 | 0.88 ± 0.45 | ||||
Autumn | 34.9 ± 0.1 | 16.2 ± 2.7 | 1.3 ± 0.7 | 0.33 ± 0.23 | 1.11 ± 0.64 | ||||
GM | Winter | 32.3 ± 1.0 | 9.7 ± 0.4 | 8.0 ± 5.3 | 1.41 ± 0.76 | 0.63 ± 0.30 | 812 | 544 | meso |
Spring | 32.5 ± 1.0 | 12.4 ± 2.8 | 2.4 ± 1.5 | 0.60 ± 0.33 | 3.03 ± 2.11 | ||||
Summer | 34.4 ± 0.4 | 18.6 ± 0.9 | 1.0 ± 0.6 | 0.66 ± 0.25 | 1.16 ± 0.72 | ||||
Autumn | 34.9 ± 0.4 | 15.3 ± 2.5 | 1.0 ± 0.6 | 1.65 ± 1.02 | 0.67 ± 0.33 | ||||
AC | Winter | 32.0 ± 0.8 | 10.8 ± 0.6 | 3.8 ± 1.1 | 1.91 ± 1.16 | 1.89 ± 1.36 | 56 | 3856 | meso |
Spring | 32.5 ± 0.6 | 13.9 ± 2.2 | 3.2 ± 1.3 | 1.25 ± 0.81 | 2.73 ± 1.23 | ||||
Summer | 33.8 ± 0.6 | 20.8 ± 0.9 | 1.3 ± 0.5 | 1.07 ± 0.69 | 2.06 ± 0.82 | ||||
Autumn | 34.3 ± 0.3 | 16.9 ± 3.1 | 2.0 ± 0.4 | 1.17 ± 0.63 | 2.24 ± 1.11 | ||||
TH | Winter | 37.9 ± 0.6 | 8.6 ± 1.2 | 1.8 ± 0.5 | 0.22 ± 0.24 | 2.42 ± 6.31 | 1295 | 125 | micro |
Spring | 36.5 ± 0.3 | 15.5 ± 3.7 | 1.2 ± 0.1 | 0.15 ± 0.14 | 0.93 ± 1.25 | ||||
Summer | 39.0 ± 1.5 | 24.9 ± 1.0 | 1.0 ± 0.2 | 0.17 ± 0.16 | 1.22 ± 0.81 | ||||
Autumn | 40.1 ± 0.8 | 17.8 ± 4.7 | 1.3 ± 0.3 | 0.32 ± 0.32 | 1.45 ± 3.38 | ||||
* Surface area calculated from the latest available map by delineating the surface of the meadow as accurately as possible at the site scale (DN: 2019 ; GM: 2002 ; AC: 2016 for ZM and 2019 for ZN ; TH: 2016).
Zostera spp. structural and morphological parameters
In order to estimate leaf cover in vegetated areas (% Cov), six quadrats (0.16 m2) were randomly placed along transects in each seagrass meadow station, and visual estimations were carried out by SCUBA divers for Z. marina and fieldworkers at low tide for Z. noltei (and Z. marina in DN). In TH, a PVC sheet was used to flatten the leaves of Z. noltei and obtain the same observation conditions as in other sites which were not submerged during observation (due to overlapping leaves). For Z. marina, all shoots in the quadrats (or a sub-sample in case of high coverage) were counted to measure the density per square meter (Dshoot). Seagrass morphometric parameters were obtained from three random PVC cores (0.005 m2 for Z. noltei, 0.03 m2 for Z. marina) sampled in the quadrats. The shoots were carefully washed free of epiphytes and sediment with filtered seawater. Fully mature leaves were measured for five random shoots per replicate. The leaf area index (LAI) was calculated as the mean leaf surface per shoot × shoot density. For Z. noltei, the density (Dshoot) was estimated from the number of shoots per core. The mean number of leaves per shoot was expressed as Nleaves.shoot and the maximum leaf height as Hleaf.max. Finally, total above-ground (leaves: Bleaf) and below-ground (roots and rhizomes: Broots) biomass was estimated for each core after freeze-drying, and the ratio Bleaf /Broots calculated to assess the relative investment of Zostera spp. between its above-ground and below-ground parts. The values (per quadrat (N=18) and per core (N=9)) were averaged to estimate all the variables at the level of the sampling point for each species (Site × Season × Modality).
Data analysis
Community Trajectory Analysis (CTA ; De Cáceres et al., 2019; Sturbois et al., 2021) was used to describe and compare the temporal trajectories of the seagrass meadows over the study seasons. CTA is based on the dissimilarity between pairs of observations in space and time (De Cáceres et al. 2019). We defined the multivariate space of resemblance between meadows using all structural and morphological traits, that were standardized before computing Euclidean distances. A Principal Coordinates Analysis (PCoA) was then used to compute the CTA metrics without any distortions (De Cáceres et al., 2019; Sturbois et al., 2021). A set of geometrical metrics was calculated to describe each meadow trajectory. The standard deviation of angles (change in direction (in degrees) from one segment to the other) measures the consistency/variability of the angles throughout the trajectory. It is complementary to the directionality metric which provides information about the consistency with which the trajectory follows the same direction, and varies between 0 and 1, with 1 corresponding to a completely directional pathway. Trajectory length represents the sum of the length of all trajectory segments (i.e. the distance between two consecutive surveys). The net change evaluates the difference between the community at the end of the study period and its initial state. The net change ratio (NCR) is defined as the ratio between the net change and the trajectory length. A low NCR indicates a decoupling between the trajectory length and the net change, so that the trajectory path does not induce as much net change as expected from segment lengths. Inversely, a high NCR illustrates that the trajectory path contributes to net changes. In this sense, NCR is complementary with directionality: if high directionally leads to high NCR, low directionality may not be equal to low NCR depending on segment lengths (Sturbois et al., 2021). Note that if trajectories are usually represented on the first two axes of the ordination (here a principal component analysis: PCA), these CTA metrics are computed using all dimensions. Finally, the dissimilarity between pairs of trajectories was quantified using the symmetrized directed segment path dissimilarity (DSDP) (De Cáceres et al. 2019) which takes into account the shape, size, direction and position of the trajectories. DSDSP was computed for both raw and centered trajectories. The dissimilarity between centered trajectories makes it possible to eliminate spatial variations that are independent of temporal trajectories (i.e. initial site-specific differences in seagrass characteristics that are time invariant). High DSDP values indicate high dissimilarities between the trajectories, while low values indicate similar trajectories.
In this analysis, both the stable and dynamic modalities of the meadow were included (except for AC, see above) allowing inter-site comparison of the two modalities. Due to missing data for Z. marina in GM and TH for winter 2020, this season was discarded for this species.
The multivariate correlations between Zostera spp. morphometric traits and environmental variables were explored by Redundancy Analysis (RDA) on standardized data. Only seagrass data from the stable modality were considered to allow simultaneous analysis of the four sites. The initial environmental data were those measured at each site during the course of the study (water temperature, light intensity, organic matter in sediment, salinity, chl-a, ammonium, phosphate), plus turbidity extracted from the REPHY network (Table 1). A stepwise selection based on adjusted coefficient of multiple determination (R2adj) was applied to the explanatory variables (Blanchet et al., 2008), and the final model selected contained only predictors with a variance inflation factor < 5.
All analyses were conducted with the R programming language version 4.3.1 (R core team 2021) and packages ‘ecotraj’ (De Cáceres et al. 2019, Sturbois et al. 2021) and ‘vegan’ (Oksanen et al., 2022).
Results
Environmental conditions
The environmental dataset can be found in Lacoste et al. (2023) and its description in Lacoste et al. (2024). Briefly, TH was characterized by a higher mean salinity over the year (39.1) and the highest and lowest water temperature values (min. 7.3°C in winter and max 26.7°C in summer). AC had a higher mean ammonium concentration throughout the year (2.8 µmol.l-1) and turbidity was also higher on average (2.1 FNU). High turbidity was also observed in GM in the winter (7.9 FNU), with an average of 1.6 FNU for other seasons. Mean light intensity in the water near the Z. marina beds was higher in DN and TH and lower in AC, whereas the Z. noltei beds were exposed to more light intensity on average in AC and TH compared to DN and GM (Fig. 2). Taking into account the periods of emersion, the light intensity in DN was as high as in AC (Fig. 2).
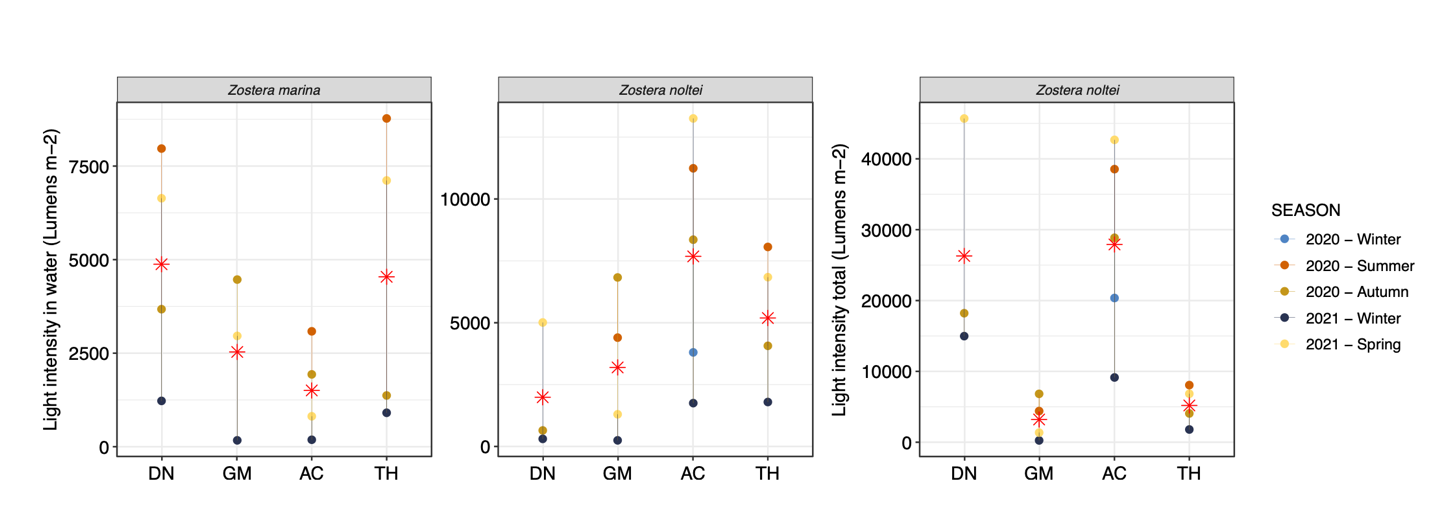
Figure 2 - Seasonal variations of mean daily light intensity at immersion (light intensity in water, lumens.m-2) for both Z. marina and Z.noltei, and at both immersion and emersion (light intensity total, lumens.m-2) for Z. noltei measured during the study near seagrass beds at each site. Red stars represent the average for all seasons per site.
Different trajectory patterns in the stable modality across sites
The two first axes of the PCA and the CTA metrics point towards site-specific seagrass trajectories for the two species (Fig. 3).
For Z. marina, AC had a long trajectory path and high standard deviation angles, but a low NCR and low directionality (Fig. 3A), meaning a pronounced variability between seasons with a return to the initial state. In contrast, the trajectory length was low in TH with a low standard deviation angle illustrating few changes from the starting point. The directionality and NCR were higher, suggesting a constant trajectory and a relative stability of ecological dynamic drivers. DN had a similar NCR and directionality to that of TH, but a much longer trajectory, indicating comparatively more changes in morphometric parameters over the study period.
When comparing trajectories (Fig. 3A), AC had the most distinct trajectory compared to the other sites, with the highest dissimilarity to DN. Although weaker, the dissimilarity remained when focusing only on the temporal patterns over the study period (i.e. on the centered trajectory; Fig. 4A2), indicating that the differences were not only due to initial spatial differences. Differences in trajectory positioning within the PCA space were mainly explained by the highest Dshoot in DN with high Broot, opposite to the more developed leaves and lower densities in AC (Fig. 5). The short length of the TH trajectory was illustrated by the low seasonal variations of the measured morphometric traits (Fig. 5).
For Z. noltei, a similar trajectory to that of Z. marina was observed in AC, albeit with a greater departure from the initial state, as indicated by similar NCR values but a longer trajectory length (Fig. 3B). As for Z. marina, TH had the shortest trajectory length and the highest NCR, indicating that the final state of this meadow results from punctual rather than gradual state changes, as illustrated by low directionality. The high standard deviation angles further suggests saltatory dynamics across the seasons (Fig. 3B). DN and GM showed more gradual changes with intermediates values of the trajectory metrics.
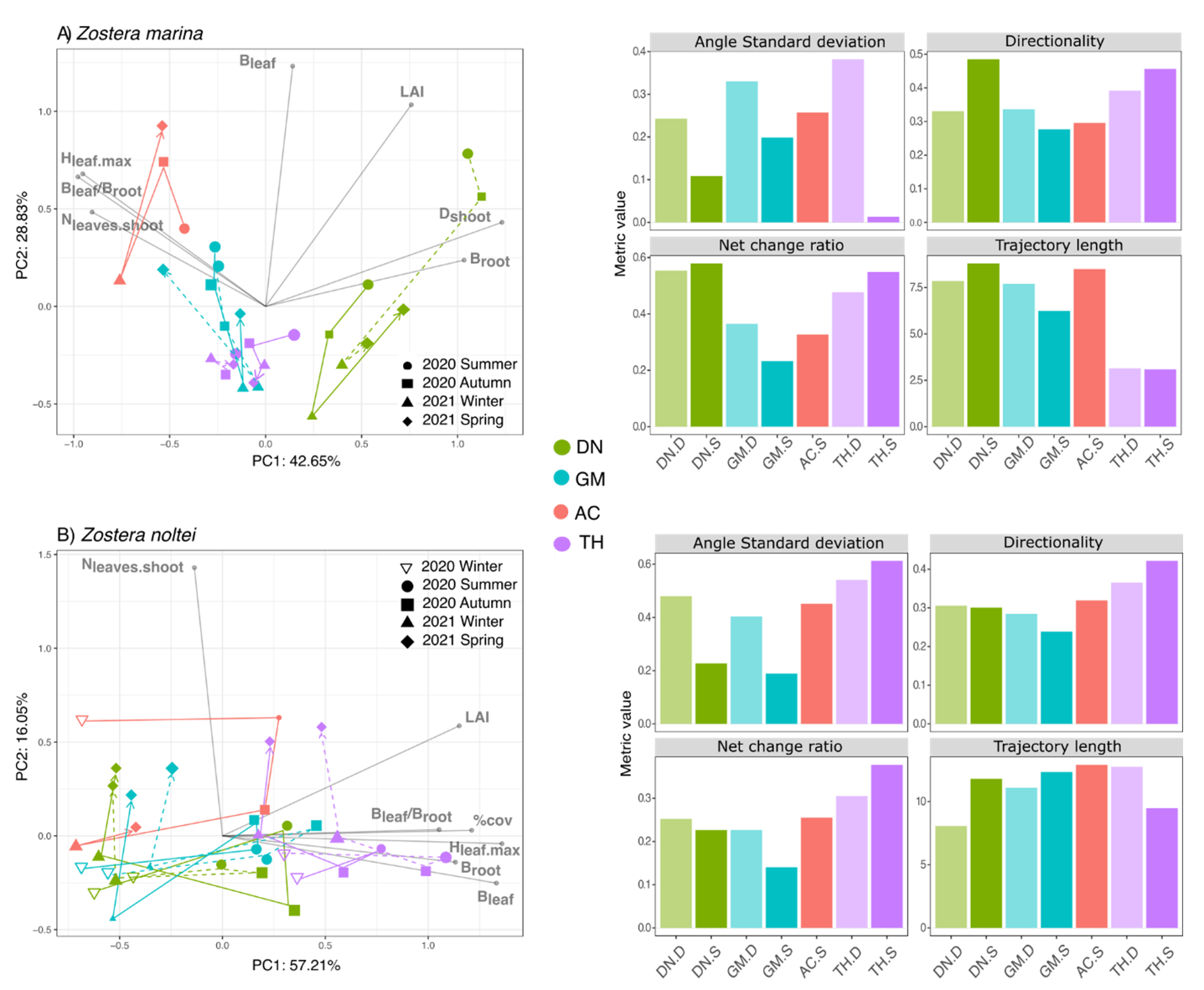
Figure 3 - Trajectory analysis of the biometric variables for A) Z. marina (summer 2020 to spring 2021) and B) Z. noltei (winter 2020 to spring 2021). The trajectories are represented in two dimensions using PCA (left panels), and trajectory metrics (using all dimensions of a PCoA with Euclidean distance) summarize the main properties of the trajectories (right panels). Biometric variables were standardized prior to both PCA and PCoA analyses. D: Dynamic (dotted lines, light bars), S: Stable (solid lines, dark bars), DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau. Samples are represented in scaling 1 while biometric variables are represented in scaling 2. The first two axes of the PCA represent ~70% of the variance for Z. marina and Z. noltei. See fig.S1 for the dissociated figure (easier to read).
TH had the most dissimilar trajectory regarding both raw and centered data (Fig. 4B). At this site, Z. noltei showed a more constant % cover over the seasons, the biomass (both Bleaf and Broot) was more important and leaves were almost twice as long as on the other sites (Fig. 6). At the three other sites, the meadows showed a clear seasonal pattern with higher % cover and shoot development in summer and autumn, whereas % cover was almost zero in winter (Fig. 6).
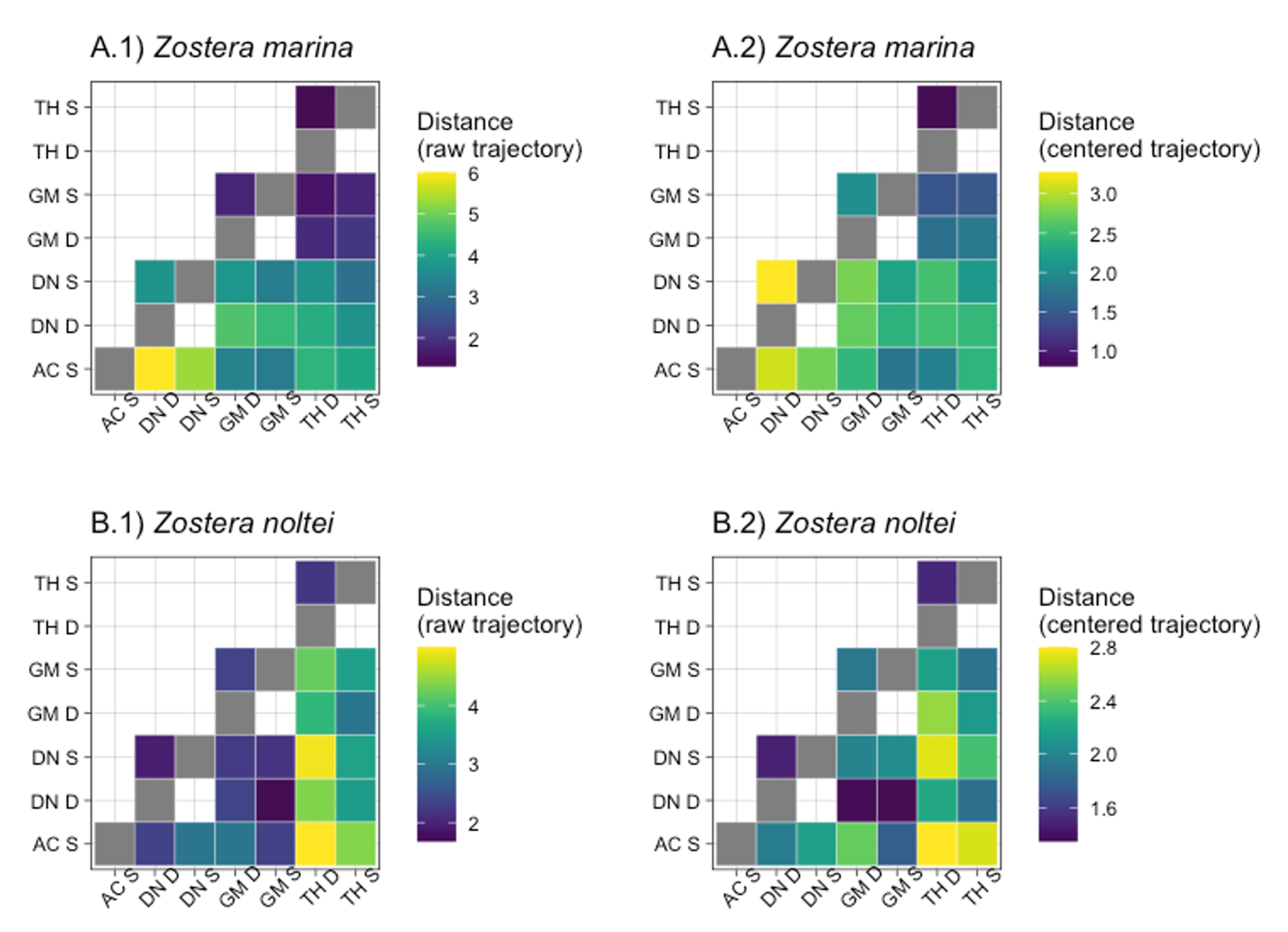
Figure 4 - Representation of the resemblance between pairs of trajectories (assessed using the symmetrized directed segment path dissimilarity; DSDSP) for A) Z. marina and B) Z. noltei. DSDSP values were computed between trajectories of the Stable (S) and Dynamic (D) parts of each meadow (DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau) for both 1) raw trajectories and 2) centered trajectories. Note that DSDSP values are not represented on the same scale for raw and centered trajectories. The higher the DSDSP values (distance), the more dissimilar the trajectories; the lower the values, the more similar they are.
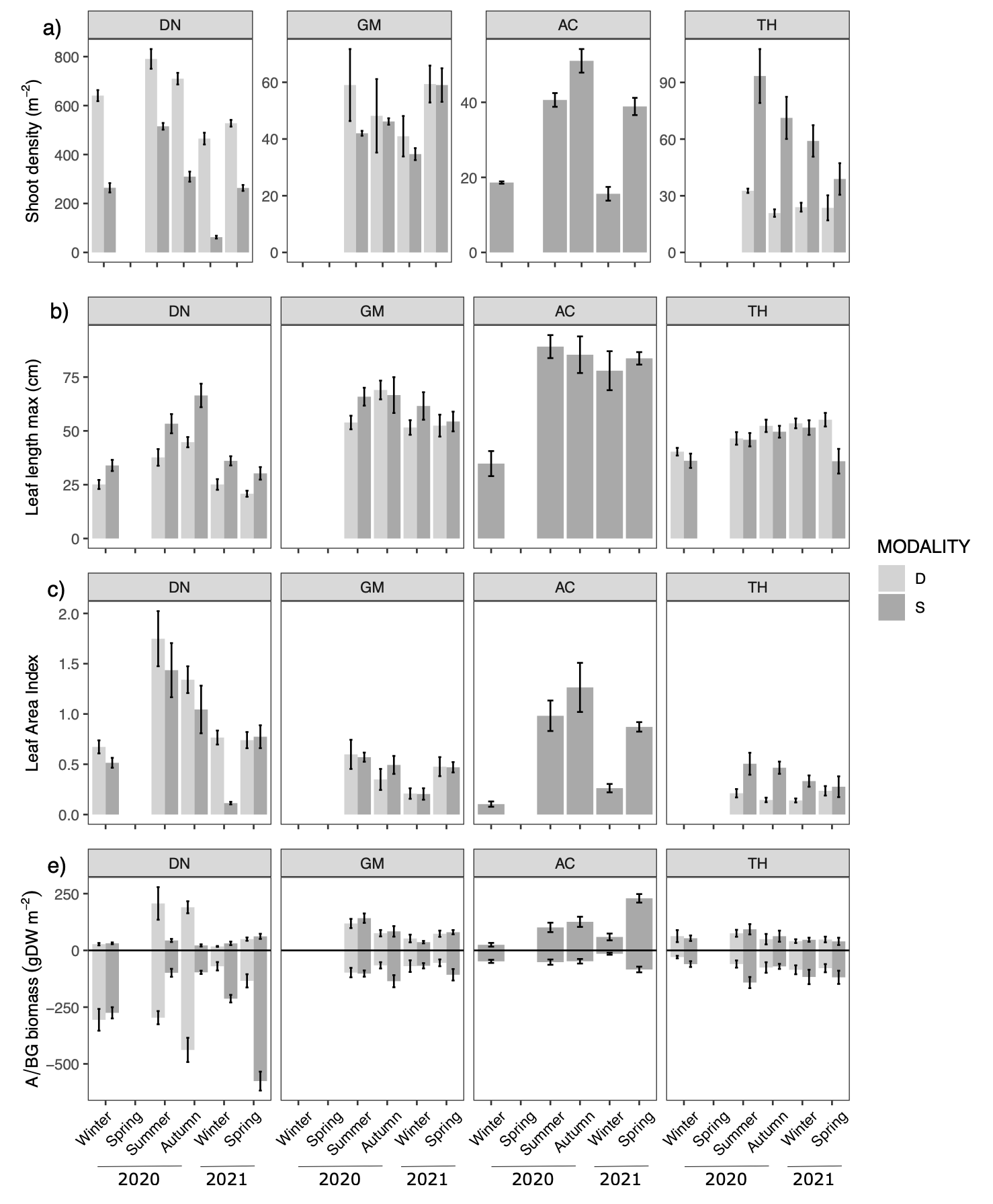
Figure 5 - Seasonal changes (mean ± se) of population descriptor: a) shoot density per m2 (note the different y-axis scales between sites), morphological traits: b) leaf length in cm, c) leaf area index, and biomass: e) above- (upper part - A) and below-ground (lower part - BG) in gDW m-2 of Z. marina at the four sites (DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau), for the two modalities (D=dynamic in light grey ; S=stable in dark grey).
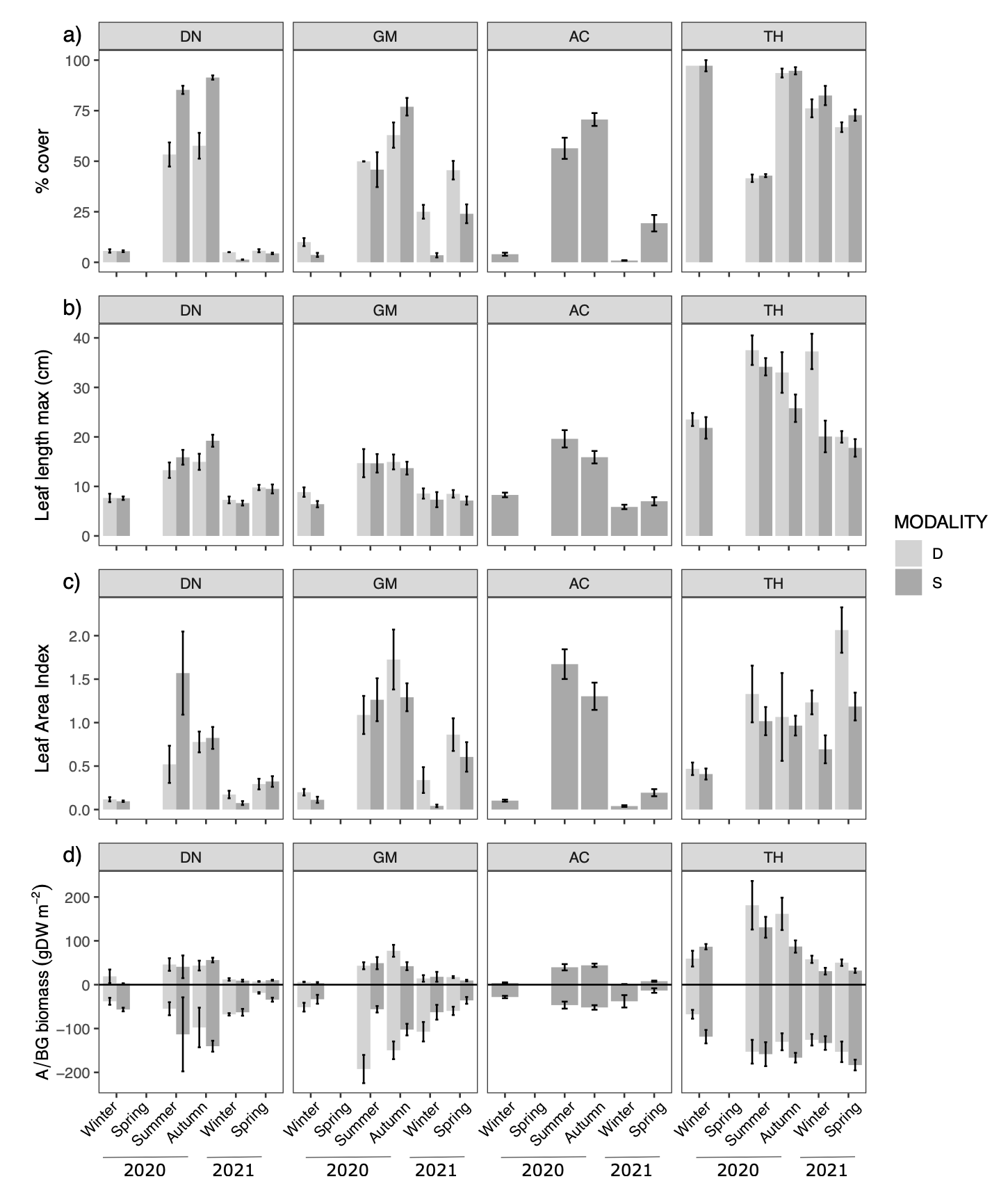
Figure 6 - Seasonal changes (mean ± se) of population descriptor: a) % cover of shoot, morphological traits: b) leaf length in cm, c) leaf area index, and biomass: e) above- (upper part) and below-ground (lower part) in gDW m-2 of Z. noltei at the four sites (DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau) for the two modalities (D=dynamic in light grey ; S=stable in dark grey).
Correlations between stable meadows and environmental conditions
The set of explanatory variables derived from stepwise selection (Fig. 7) accounted for a differing amounts of morphometric trait variation depending on the species, with 32% and 56% (p < 0.05) for Z. marina and Z. noltei respectively. The explained variation matched to some extent with the main patterns emphasized by the unconstrained ordination of Zostera spp. traits (Fig. 3), discriminating sites.
Higher shoot densities and below-ground biomass in Z. marina were associated with higher mean light intensity, as observed in DN (Fig. 7A). In contrast, greater leaf size was associated with a lower intensity of light received, and lower salinity and PO43- concentrations, which corresponds to the conditions prevailing in AC (Fig. 7A).
High biomass, % cover and leaf size of Z. noltei were associated with high salinity and low nutrient concentrations (Fig. 7B) that characterise the environmental conditions of TH, where the meadows are also permanently submerged. Leaf development (Nleaves.shoot and ratio Bleaf/Broot) was positively correlated with mean light intensity received by plants during immersion.
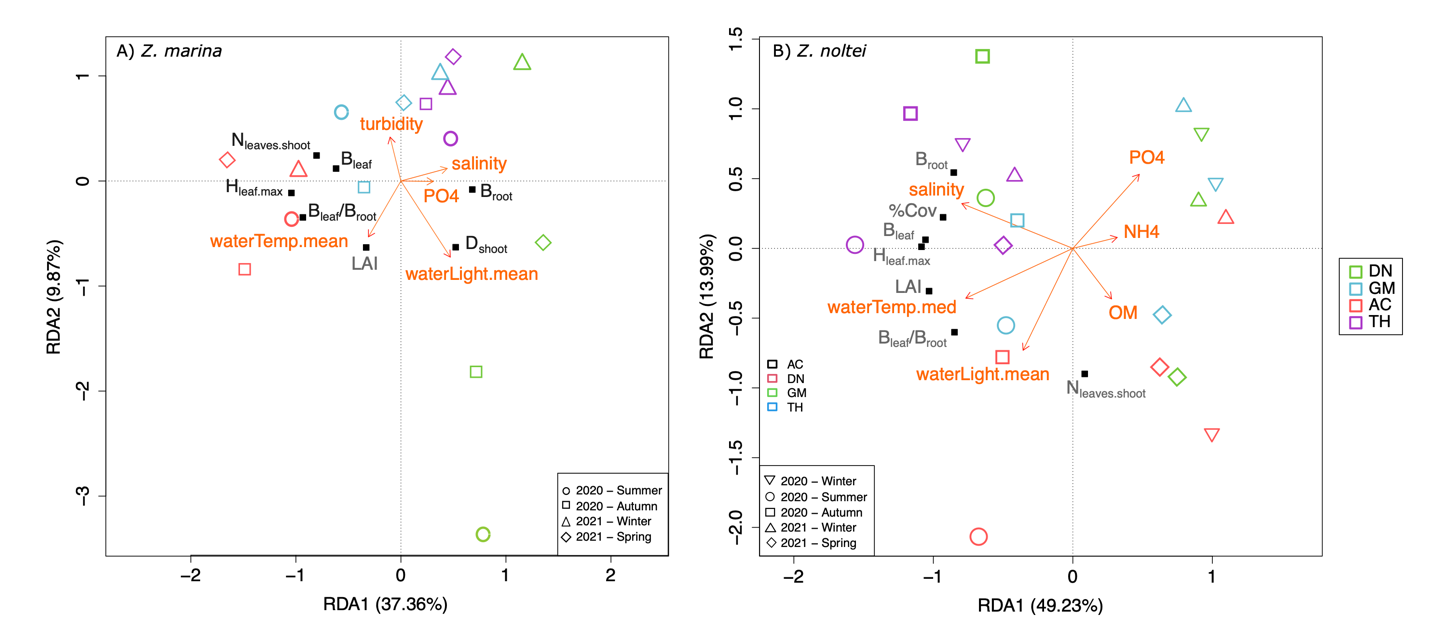
Figure 7 - Redundancy analysis of the standardized morphometric traits and population descriptors (in black) of A) Z. marina and B) Z. noltei against selected environmental variables (in orange). Within-site dispersion represents the seasonal variability. Scaling type 2. (DN: Dinard, GM: Morbihan, AC: Arcachon, TH: Thau). Bleaf and Broot: leaf and below-ground biomass, Hleaf.max; max leaf length (cm), Dshoot: shoot density, LAI: leaf area index, %cov: percentage cover , Nleaves.shoot: number of leaves per shoot.
Local variability of the seagrass traits
In Z. marina, the variation of the standard deviation angle was always higher in the dynamic modality of the meadows (Fig. 3A). The main differences between the trajectories of the stable and dynamic modalities were observed in DN (Fig. 4A). Regardless of the season, DN showed a higher Dshoot within the dynamic modality, characterised by less developed leaves (Fig. 5). In summer and autumn, biomass in DN was also largely higher in the dynamic modality than in the stable one (Fig. 5). A contrasting observation was made for TH, where a consistently higher Dshoot was observed in the stable modality compared to the dynamic modality (Fig. 5).
The Z. noltei habitat appeared to be more homogeneous, with less dissimilarity between the trajectories of the two modalities at the three sites (Fig. 4B), although a higher standard deviation of angles was observed in the dynamic modality at GM and DN (Fig. 3B). The most consistent pattern of difference was the presence of a higher Broot within the dynamic modality of GM, with also a higher % cover compared to the stable modality during winter and spring (Fig. 6). In DN, differences in % cover between the stable and dynamic modalities were observed in summer and autumn when the recovery percentage exceeds 50%. (Fig. 6).
For both Zostera species, inter-site variation was more important when comparing samples from the dynamic modality than when considering the stable meadows (Fig. 4).
Discussion
Although the seasonal dynamics of Zostera species are well documented (e.g. Azcárate-García et al., 2022; Cognat et al., 2018; Ouisse et al., 2011, 2010), few studies have compared these dynamics at regional and local scales, including several sites and different positions within seagrass beds. Through an extensive fieldwork, this study allowed us to identify several temporal dynamic patterns of Z. noltei and Z. marina, and to correlate different population and phenotypic strategies to environmental variations.
Regional scale Zostera spp. dynamics
The seasonal pattern of Z. marina observed in this study was consistent with the description of other populations distributed at equivalent latitudes, where temperature and light favour plant development in summer, while metabolism is reduced in winter (Carstensen et al., 2016; Lee et al., 2007). The exception was the TH site where the trajectory during the study period was very short, highlighting the low variation in plant morphological traits throughout the year. At this site, the permanent subtidal nature of this population, exposed to low currents and no air, is likely to limit the winter decline of the population observed at other sites.
Unlike intertidal populations from southern Spain, which have been shown to potentially reach their physiological limits during summer (Azcárate-García et al., 2022), no negative effect of high summer water temperatures was observed on the TH population during the study period. This suggests that the subtidal location of this population may be advantageous compared to intertidal populations in the southernmost range. However, the potential for Z. marina to adapt to increasing water temperatures at this particular site remains to be described, particularly in the case of abrupt temperature changes such as during heat waves. In addition, such a subtidal strategy may also pose some challenges, particularly in terms of access to light, which may be limited by depth and eutrophication (Krause-Jensen et al., 2021).
Seagrass meadows have been shown to exhibit diverse strategies across different study sites, reflecting the significant influence of environmental conditions (Azcárate-García et al., 2022; Soissons et al., 2018b). Opposite strategies were illustrated in our study for two populations of Z. marina, which differed through their investment in shoot density vs leaf size, according to environmental constraints. The population in DN, which is exposed to strong tidal currents, invested in shoot production and below-ground biomass, which is particularly suited to resist erosive forces (Peralta et al., 2008). This population, with its smaller leaves, seems to correspond to the intertidal strategy of Z. marina (angustifolia) reported by (den Hartog, 1972) and later by Becheler et al. (2010) in Brittany (St Malo), adapted to more stressful environmental conditions. In contrast, in AC, a more sheltered area with lower light exposure, an opposite strategy was observed with a more important investment in leaves, which is typical of light-limited meadows (e.g. deep meadows), maximizing their photosynthetic parts (Krause-Jensen et al., 2000; Li et al., 2013). Such phenotypic variation has been synthesized by Boyé et al. (2022), who defined two types of trait configuration in Z. marina Brittany populations. According to their model, the DN population corresponds to type 1, exposed to stressful intertidal conditions, whereas the AC population falls into type 2, with a phenotype mostly constrained by limited light. In the absence of significant strong hydrodynamic conditions, the strategy of the GM and TH populations tends to be intermediate with no particular investment in shoot density but reduced leaf development due to less restricted access to light compared with AC. At this scale, the role of other pressures on population structure not considered in this study, such as pollution (Scarlett et al., 1999) or physical damage (e.g. boat moorings, Ouisse et al., 2020) could also contribute to explain the ecological strategy.
Difference in the trajectories of Z. noltei between sites was mainly driven by the constantly higher % cover observed in TH, largely due to the specific environmental conditions of this microtidal site (permanent immersion). The absence of tidal currents at this site likely promoted leaf development, and increased the ratio of above- to below-ground biomass, whereas it was reduced at the other sites with tidal current. Like Z. marina, Z. noltei at the TH site maintained its biomass during winter and showed a persistent cover, which may be explained by the more stable hydrological conditions i.e. no mechanical forcing due to tidal current. At the other intertidal sites, the seasonal pattern corresponds to the annual strategy of this species, which loses most of its aboveground biomass during winter due to harsh conditions. Another important factor that may explain the drastic decline in density during winter is the predation by herbivorous birds, which has been shown to cause significant regression of seagrass beds in DN and GM (Gaudard et al., 2017).
Local scale Zostera spp. strategies
Variability in population and individual characteristics at the patch scale may result from spatial differences in mortality, recruitment and production, under the effect of hydrodynamic control and/or human activities (Brun et al., 2003; Duarte and Sand-Jensen, 1990). Locally, seagrass growth may also vary significantly in relation with microtopography (e.g. tidal pools) or light reduction due to macroalgal mats (Cognat et al., 2018). Balestri et al., (2003) showed variability in basic structural, morphological and dynamic characteristics used as indicators of the status of Posidonia oceanica at a spatial scale smaller than metres. In our study, the most obvious within site differences were observed for Z. marina in DN, where the shoot density was higher in the dynamic area while the leaves were smaller. This could be the result of stronger tidal currents at the dynamic position, where the meadow experienced periods of emergence during the spring tide, which could reinforce the expression of the intertidal facies of Z. marina described above. Although less important, the difference in shoot density observed between the two areas in TH suggests that parameters other than hydrodynamic control fine scale Z. marina development. At this site, a higher biomass of macroalgae was observed in the dynamic area compared to the stable area (Lacoste et al. 2023), which may have resulted in competition for light and thus lower shoot development (Nelson and Lee, 2001). However, this hypothesis does not seem to be generalisable in a larger context as the observation of a higher macroalgal biomass in the stable area compared to the dynamic area in GM did not have such an effect on Z. marina. Other biological factors such as the diversity associated with seagrasses (e.g. grazer biomass), small scale hydrodynamic conditions (e.g. fetch) or local anthropogenic pressures (e.g. pollution, anchoring) may also contribute towards differences in growth.
One of the main limitations of this study was the lower resolution of the available environmental data (site level) compared to the scale of observation of seagrass beds (i.e. stable vs dynamic modality). This limits our ability to explain the site-specific dynamics in the different parts of the meadow, which may be due to a combination of unmeasured factors.
Although the causes of the observed local variability require further investigation, the results of this study confirmed that there can be differences in the dynamics of Zostera spp. beds at the scale of hundreds of meters. Most importantly, our results showed that such local variability can lead to differences in interpretation when comparing the ecological trajectory of seagrasses at broader scales. This highlights the importance of collecting as much information as possible on environmental drivers but also on anthropogenic pressures, at the appropriate scale, so as not to confound local adaptation with more global drivers of species’ ecological strategy, including human impacts. This also has implications for monitoring strategies and management actions, highlighting the need for in-depth knowledge of species to better design monitoring and management actions.
Perspectives for research and monitoring
Given the different ecological strategies adopted by Zostera species, and by populations within species, the question is how these different strategies will affect the resilience of populations in the coming years under changing environmental conditions and cumulative pressures. Photosynthesis is expected to be less sensitive to heat in high light conditions (Berry and Bjorkman, 1980), which could for example give the Z. marina DN population an advantage over the AC population. Rising temperatures are also likely to increase the productivity of the northernmost populations (Beca-Carretero et al., 2018; Cott et al., 2021). However, higher temperatures has also been shown to reduce the shoot density in Z. marina (Richardson et al., 2018), which may be incompatible with the strategy of shoot development in highly hydrodynamic conditions, such as in DN (Boyé et al., 2022). In Thau lagoon, the subtidal conditions appeared to suit the development of a perennial Z. noltei population. Mediterranean coastal lagoons are however among the ecosystems most threatened by climate change (Lloret et al., 2008), and the predicted increases in temperature coupled with heatwave intensification (Cramer et al., 2018) could threaten their conservation. Z. noltei Atlantic populations have been shown to be severely affected by warming conditions (Repolho et al., 2017) and higher temperatures have also been shown to increase the sensitivity of Z. noltei to some contaminants (Gamain et al., 2018). Despite this threat, the impact of marine heatwaves on Z. noltei in the Mediterranean has not yet been studied. More largely, the response of seagrasses to cumulative pressures is still largely unpredictable, ranging from negative (negatively synergistic or additive) to positive (positively synergistic or antagonistic) (Stockbridge et al., 2020), with varying degrees of intensity.
Because of their widespread distribution and sensitivity to a wide range of human-induced disturbances, seagrasses are used as biological indicators in several monitoring programs. Highly sensitive to water clarity and eutrophication (Dunic et al., 2021), they are considered to be particularly good indicators of water quality for the European Water Framework Directive (WFD, 2006/60/EC, e.g. Marbà et al., 2013). In this context, Z. marina and Z. noltei are jointly assessed (together with other macrophytes for the Mediterranean coasts of France, Italy and Greece) to establish an angiosperm (in the Atlantic and English Channel) or a macrophyte (in the Mediterranean) indicator, with a reference state defined at a large scale. In line with others, our study suggest however the high adaptive capacities of Zostera spp. to regional and local environmental conditions, which induce variability in population and morphometric traits at varying scales. The temporal trajectories were also site-specific and habitat characteristics varied according to the sampling period. Thus, there is a risk of overgeneralisation in the interpretation of large spatial scale indicators in heterogeneous habitats if the sampling design does not adequately account for i) within-site variability resulting from small-scale patchiness, as shown here at the modality scale, and ii) temporal variation at the appropriate scale. At a time when authors are increasingly advocating ecological study scale expansions for both marine conservation and theoretical purposes (Edgar et al., 2016; Witman et al., 2015), we also recommend the multiplication of sampling stations at the local scale and the replication of sampling in time for an accurate assessment of the state of coastal waters. Non-destructive sampling is also strongly recommended to avoid further pressure on natural populations.
Data, scripts, code, and supplementary information availability
Data and supplementary information are available in Lacoste et al. 2023: https://doi.org/10.17882/97962. The script can be found at: https://doi.org/10.5281/zenodo.10427793
Acknowledgments
This study is part of the Life Marha project funded by the European Union. The authors would like to thank Lionel Allano, Aurélie Foveau, Aurore Lejolivet, Titouan Brébant, Marine Pasquier and Pierre-Olivier Liabot for their help in the field and/or in the laboratory.
Preprint version 3 of this article has been peer-reviewed and recommended by Peer Community In Ecology (https://doi.org/10.24072/pci.ecology.100638; Vernay, 2025)
Funding
The authors declare that they have received no specific funding for this study.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.