
 CC-BY 4.0
CC-BY 4.0
Introduction
The demand for forest biomass has been increasing in Europe because forest biomass can be used for wood products, paper production, and even for energy production (Nicholls et al., 2009; Diaz-Yanez et al., 2013; Achat et al., 2015a). An approach to meet this growing demand is to intensify forest management, for instance planting dense stands of fast-growing species managed in short-rotations (Ranger & Nys, 1996; Heilman & Norby, 1998; Aylott et al., 2008; Achat et al., 2018). However, intensifying forestry has shown to be potentially deleterious for ecosystem functioning such as impoverishing soils (Kimmins, 1974; Achat et al., 2015a) and such an approach needs to be evaluated (Toman & Ashton, 1996; Fox, 2000; Pohjanmies et al., 2021). Evaluating intensive forestry is all the more necessary because this kind of forest management exports large quantities of nutrients, whereas natural nutrient inputs are generally of low magnitude (Kimmins, 1974; Heilman & Norby, 1998; Ranger & Turpault, 1999; Flueck, 2009; Achat et al., 2015a). In addition, the impacts of management on forest carbon and water balance and its biophysical functioning is also observed at various spatial scales (Magnani et al., 2007; Albaugh et al., 2016; Luyssaert et al., 2018). Therefore, the main objective of the present study was to assess the impact of different levels of forestry intensification on ecosystem functioning. In practice, we evaluated the effects of three forest management alternatives (FMA), a theoretical framework developed to compare contrasting silvicultural systems (Mason & Meredieu, 2011). The studied FMA manipulated stand density, competition control, fertilisation, genetics, and soil preparation, so they fulfil all the criteria of the intensive management definition (Fox, 2000).
One major difficulty when assessing forestry strategies is the time dimension because forest management has to be evaluated over decades, if not centuries (Homann et al., 2000; Kimmins et al., 2008). To circumvent this obstacle, different approaches are possible such as modelling (Wallman et al., 2005) or long-term in situ monitoring (Pretzsch et al., 2019). While we value models and regularly use them (Loustau et al., 2005; Jonard et al., 2010; Achat et al., 2018; Moreaux et al., 2020), the present study relies on a long-term experimental platform. In addition to being crucial for improving ecosystem models (Achat et al., 2016a), long-term field experiments enable accurate assessments of ecosystem responses to silviculture (Prescott, 2014) and are useful for understanding complex ecosystem functioning (Green et al., 2023). Consequently, we set up a long-term forestry experiment in 2012 in south-western France, a region characterised by debates regarding intensive versus extensive forest management scenarios (González-García et al., 2014; Mora et al., 2014) and we monitored the ecosystems under different silvicultural systems. Hereafter, we report the results of one decade of monitoring that showed how intensively managed forest plantations have influenced biogeochemical and biophysical aspects of ecosystem functioning (Fox, 2000). Based on the literature, we had three main expectations at the beginning of the monitoring: one about biophysical effects of FMA and two about biogeochemical effects. Firstly, we expected that contrasting stand densities would have short-term effects on the ecosystem biophysical functioning (Pohjanmies et al., 2021) as a time lag in canopy closure can influence the local microclimate. More precisely, our hypothesis (H1) was that dense plantations would induce cooler and drier conditions than in sparse stands (De Frenne et al., 2021; Zhai et al., 2024; Chen et al., 2025) because of high sunlight interception, rainfall interception, and evapotranspiration (Ewers et al., 2005; Barbier et al., 2009; De Frenne et al., 2021). Secondly, because ecosystem biogeochemical functioning is mainly influenced by forestry in mature stands (Erisman & Draaijers, 2003; Johnson & Turner, 2014), or through major disturbances (Weis et al., 2006; Smolander & Heiskanen, 2007; Paré & Thiffault, 2016), we anticipated that the young forest stands would not display any strong biogeochemical differences between FMA (H2). Finally, we identified an exception to this H2 expectation, which was the introduction of N-fixer plants, because this practice showed short-term modifications of the ecosystem N cycling (Dyck et al., 1983; Watt et al., 2003; Bouillet et al., 2008; Voigtlaender et al., 2012; Vidal et al., 2019, 2021). We consequently hypothesised an increase of nitrate production and an improvement of tree N nutrition under such conditions (H3).
Material and Methods
Study region
The XyloSylve forest experimental platform was installed in 2012 in the Landes de Gascogne region (∼43.5-45.5°N and ∼1.5°W and 0.3°E), which is located in south-western France (Figure S1). The regional climate belongs to the temperate, fully humid, warm summer class (Cfb) of the Köppen-Geiger classification (Kottek et al., 2006). The local climate is monitored in an open area near (<1 km) the forest stands and data from the last two decades indicate mean annual values of 13.1°C for air temperature and 908 mm yr-1 for bulk precipitation. Summer precipitation (May-September) is on average 292 mm yr-1, resulting in moderate water stress (monthly SPEI values=4.7; Hargreaves climatic moisture deficit=354 mm yr-1; Figure S2). Nevertheless, climatic indices varied a lot from year to year during the period studied (Table S1), implying possible droughts or heatwaves.
The Landes de Gascogne region is a 1.3 Mha flat area, characterised by a low elevation and a sparse network of rivers (Jolivet et al., 2007; Deirmendjian et al., 2019). Due to these properties, the water table level fluctuates near the soil surface (0-3 m deep), except near the rivers that function as natural drainage ditches (Cottinet, 1974). The water table is characterised by oligotrophic conditions but levels of redox and oxygen concentration vary over time (Righi, 1977; Jolivet et al., 2007; Deirmendjian et al., 2019). The soil parent material consists of Aeolian deposits of quaternary sands (∼ 23-15 kyr BP), which are relatively homogeneous in particle size (mainly 100-200 µm) and rich in quartz (Juste, 1965; Bertran et al., 2011; Sitzia, 2014). Aeolian deposits of this kind are found in several regions of Europe, such as in England, Netherlands, Denmark, Germany and Poland (Sitzia, 2014). While they are dominated by the quartz fraction, these sands also contain substantial amounts of orthose, but very low contents of phyllosilicates or minor minerals such as tourmaline, staurodite, zircon and andalousite (Juste, 1965; Latouche, 1971; Righi & De Conninck, 1977). Soils that have developed from the Aeolian deposits are mainly non-structured sandy podzols (albic; entic and/or ortsteinic; Augusto et al., 2010). Podzols with the ortsteinic characteristic have a cemented B soil horizon –locally named alios– that physically limits the vertical distribution of plant roots and hence plant growth (Delibrias et al., 1966; Gelpe, 1992). Nevertheless, because of the discontinuous nature of this alios horizon (Richer-de-Forges et al., 2017), soil exploration by tree roots is only partly impeded by this horizon (Achat et al., 2008) whereas the presence of the water table may be a major obstacle to tree rooting in the wet areas (Bakker et al., 2006). Soils are acidic and poor in nutrients and oligo-elements (Augusto et al., 2010, 2022), such as potassium or magnesium (Delmas, 1954; Demounem, 1979). Nevertheless, it is the phosphorus (P) deficiency that characterises most these soils as the extremely low soil Ptotal content (Achat et al., 2009; Augusto et al., 2010) makes them some of the poorest soils worldwide (Lambers et al., 2010; He et al., 2021). In podzols, the soil physical-chemical reactivity is generally led by organic matter and aluminium-iron oxides (Eusterhues et al., 2005; Jansen et al., 2005; Grand & Lavkulich, 2013, 2015; Achat et al., 2016b), a characteristic that applies to the soils of this region (Augusto et al., 2010; Achat et al., 2011).
Study forest
Created in the second half of the 19th century (Jolivet et al., 2007), the Landes de Gascogne forest covers 0.9-1.0 Mha, making it the largest artificial forest in Europe (Figure S1A). Mainly composed of even-aged stands of maritime pines (Pinus pinaster Ait.), this forest is managed for wood production, pulp and industrial fuelwood (Jolivet et al., 2007; Orazio et al., 2015). Forest management includes plant breeding, soil preparation at planting, drainage, P-fertilisation, vegetation control, and several thinnings before a final clear-cut at 35-50 years-old (Dorlanne, 1991; Chaperon & Cremiere, 1994). Such intensive management, in conjunction with the adaptation of the maritime pine species to harsh conditions (Chevalier et al., 2024), enable a stand productivity that is moderate in comparison to European forests (Tóth et al., 2013), but that is high when taking the local soil poverty into account.
It is worth mentioning that the Landes de Gascogne forest is exposed to major disturbances (Senf & Seidl, 2021), such as huge winter windthrows (in 1999 and 2009), summer droughts (in 2011, 2016 and 2022), and large-scale wildfires (in 2022) due to the high ignitability of maritime pines (Tapias et al., 2004; Fernandes & Rigolot, 2007). In addition to those hazards, the sustainability of the system is threatened by current changes in silvicultural practices. Indeed, while the harvesting intensity of this forest is among the highest in Europe (Levers et al., 2014), recent technical innovations have enabled the generalisation of rooting system harvests and –to a lesser extent– whole-tree harvests (Banos & Dehez, 2017), with possible deleterious consequences on ecosystem functioning (Achat et al., 2015a; Achat et al., 2015b; Augusto et al., 2022).
Experimental platform
The XyloSylve experimental platform was initiated with the objective of testing, under real conditions, innovative silvicultural systems that were expected to supply in biomass in the regional forest-wood chain. In a first stage, discussions were organised between scientists and stakeholders to define forest management alternatives (FMA; Mason & Meredieu, 2011) that would constitute a gradient of extensive-intensive approaches, in combination with a conservative-innovative gradient. A list of eight silvicultural systems was finally defined for field evaluation and three were evaluated in the present study (see below). These three silvicultural systems were contrasted enough to belong to different classes of the FMA framework (Mason & Meredieu, 2011).
The XyloSylve platform was established in the INRAE experimental forest (elevation ≈ 65 m above sea level). The area belongs to the “wet moorland” type based on the regional site classification (Jolivet et al., 2007), meaning that the water table level remains fairly close to the topsoil level (i.e. 0–2 m deep), and that spontaneous vegetation is dominated by purple moor-grass (Molinia caerulea) and to a lesser extent by bracken (Pteridium aquilinum) or common heather (Calluna vulgaris). The XyloSylve platform consists of two distinct sites about 1.5 km apart: (1) the trial site (44.7352–44.7390°N; 0.7918–0.8005°W) assesses the practical feasibility of the eight FMA, but its results are not presented in the present study; and (2) the monitoring site (44.7441–44.7505°N; 0.7718–0.7805°W), which provided data for the present study. The monitoring site is composed of three large plots (∼8–9 ha each) that test three contrasting silvicultural systems (see below). These three plots were side-by-side and were located on a flat horizontal plain, without any geomorphological differences (slope, elevation, microrelief). They were equipped with sensors and collectors to assess biogeochemical and biophysical changes, from the atmosphere to the groundwater. The large surface area of each plot was chosen to enable monitoring the carbon-water fluxes using the eddy-covariance methods (Kowalski et al., 2003; Barcza et al., 2009). The practical consequence of having large, heavily equipped, plots was that it was not feasible to test all of the eight silvicultural systems, or to replicate them. The absence of true replication implied that it was not possible to evaluate our results based on statistical tests because sampling replicates or repeated measurements over time are pseudoreplicates (Hurlbert, 1984). However, Hurlbert also concluded that “Replication is often impossible or undesirable when very large-scale systems are studied [...] or when the cost of replication is very great”, which is the case of our experiment. We followed Hurlbert’s recommendations for this kind of study, which is not to use inferential statistics, but descriptive analyses instead.
Forest management alternatives (FMA)
Foresters may influence ecosystem functioning through several practices such as tree species choice (De Schrijver et al., 2008), vegetation control (Balandier et al., 2006), or thinning regimes (del Campo et al., 2022). In the three forest management alternatives (FMA) retained for the monitoring site, we paid particular attention to stand density and management of the understory. Indeed, stand density influences productivity while strongly modifying hydrological and biogeochemical processes (Andre et al., 2008; Hall & Marchand, 2010; Cai et al., 2016). Stand density affects light transmittance (Gaudio et al., 2011; Dumas et al., 2022), so it interacts with the dynamics of the understory vegetation and the latter is the second factor taken into account in our selected FMA. It is all the more relevant because spontaneous vegetation is abundant and functionally important in the study region (Rivalland et al., 2005; Jarosz et al., 2008; Gonzalez et al., 2013; Vidal et al., 2021).
The three FMA were tested in plots formerly occupied by mature pine stands, which had been clear-cut, with biomass export during the winter of 2012-2013. The site preparation was also the same for all the FMA, including soil ploughing-disking and fertilisation with inorganic phosphorus (Table 1), a practice long-known for improving tree growth in the study region (Trichet et al., 2009). Then, FMA differed from each other (Table 1; plantations in early-spring 2013): the wood biomass production (WBP) was designed using a very intense forestry approach (FMA5 in Mason & Meredieu, 2011). It included a dense plantation (Figure S3A) of a seed lot of maritime pine that was selected to be fast-growing (Table 1). The combined objective management (COM; FMA3 in Mason & Meredieu, 2011) aimed at improving pine growth by alleviating any competition. Inter-tree competition was limited by planting at low density (Figure S3B) while competition due to spontaneous vegetation (Bon et al., 2023) was reduced with periodic mechanical controls (Table 1). This FMA will include the plantation of broadleaved species (birch and oak) dedicated to future production diversification and support for biodiversity, which explains the management name, but this is not in the scope of the present study (see § Perspectives). The nutrient management FMA (NuM) was designed to improve the regional standard silvicultural system (moderate stand density and vegetation control; Figure S3C) using a nature-based solution, namely sowing N-fixers in the stand furrows. The species used were common gorse (Ulex europaeus L.; 100 seeds m-2) and common broom (Cytisus scoparius (L.) Link; 100 seeds m-2), which are local shrubs. Common gorse proved to fix atmospheric nitrogen (N) at a high rate (Augusto et al., 2005; Vidal et al., 2019) and was expected to be the species that is the best adapted to local conditions. In general, all of the FMA were designed to produce biomass, but with a level of forestry intensity that ranks: WBP>COM>NuM.
Table 1 - Tested forest management alternatives. The forest management alternatives are fully described in the Methods section.
Characteristics | Wood biomass production (WBP) | Combined objective management (COM) | Nutrient management (NuM) |
|---|---|---|---|
Past land-use | mature maritime pine, clear-cut including stump harvest | ||
Soil preparation | ploughing and disking | ||
Fertilisation | 50 kg-P ha-1 | ||
Plantation | 2500 trees ha-1 (2×2 m) | 952 trees ha-1 (1.5×7 m) | 1250 trees ha-1 (2×4 m) |
Tree species | Pinus pinaster (“biomass” seed lot)# | Pinus pinaster | Pinus pinaster |
N-fixer sowing | none | none | Ulex europaeus |
Vegetation management | 1 control (at 8 yrs) | 5 controls (at 4-8 yrs) | 2 controls (at 5 and 8 yrs) |
Intensity of management | very high | high | moderate |
(#) The VF3 seed lot is issued from the third series of seed orchards from the French maritime pine breeding program (breeding within the SW France population, based mainly on growth and form). This improved seed lot is largely used in the Landes de Gascogne forest. Conversely, the “biomass” seed lot is an experimental (non-commercialised) mix of half sib families, the parents of which were selected from the same generation as VF3 parents, but for high growth rate (Annie Raffin, INRAE, personal communication).
Study design
The monitoring site is composed of three large plots of ∼8-9 ha, arranged side-by-side (Figure S4A). In each plot, monitoring was carried out in three subplots: two peripheral subplots (in NE and SW directions) dedicated to most measurements and one subplot dedicated to water table sampling and eddy-covariance monitoring (the central area; Figure S4B). The rationale of this design is to let undisturbed the biogeochemical monitoring (in the NW-SW peripheral subplots) when pumping the water table for sampling in the central area. Similarly, the eddy-covariance monitoring was carried out in the central subplot because this method takes into account processes over large surface areas (Kowalski et al., 2003) and consequently it was necessary to locate the eddy-covariance tower far away from any forest edge. Similarly, we located the two peripheral subplots at least 40 m away from the closest forest edge, because the vicinity of an edge (or a ditch) may affect the local biogeochemical functioning (Wuyts et al., 2008; Ågren et al., 2024). The monitoring is presented below but also summarised in Table S2.
Soils
The initial properties of soils were studied based on a large sampling campaign. In practice, each plot was studied using two complementary sampling approaches: (1) in the central hectare of the plot, a deep sampling (0–120 cm deep) was made at 36 points using a 6×6 systematic grid, designed to capture the mid-distance soil variability. This approach was designed also to produce a limited number of representative composite samples on which numerous different chemical determinations could be made (see below). (2) additional sampling points (0–60 deep) were distributed at random in each plot (N=72–88 depending on the surface area of each plot). This second sampling approach was designed to capture the spatial variability of a few properties (C, N, pH) within each plot.
Soils were sampled using a gouge auger driven into the soil with a percussion drill. The soil cores were described (presence of roots, soil horizons, induration) before being split into layers of systematic thickness (0–15 cm, 15–30 cm, 30–60 cm, 60–90 cm and 90–120 cm). For approach (1), all samples of a given layer were combined to provide five composite soil samples per plot.
The soil properties of the mineral layers were determined using standard French methods (NF): Soil pH-H2O was measured in a water/soil suspension (1:2.5; NF ISO 10390). Total C and N contents were quantified by dry combustion (NF ISO 10694 and NF ISO 13878). Total C was considered as soil organic carbon (SOC) because the studied soils contain no carbonate (Augusto et al., 2010).
Additional chemical analyses were performed, but only on composite samples (i.e. one composite sample per soil layer [0–15 cm, 15–30 cm and so on; approach (1)] and per plot). The additional analyses were as follows: for soil texture, the five-size fractions for clay (<2 μm), fine silt (2–20 μm), coarse silt (20–50 μm), fine sand (50–200 μm), and coarse sand (200–2000 μm) were assessed (NF × 31–107). Extractable P content was quantified (Olsen et al., 1954). EDTA extractions were done for Cu, Fe, Mn and Zn quantification (NF 31–120). Boron was extracted using the hot water method (NF 31–122). Total content of 17 elements (Cr, Cu, Ni, Zn, Co, Pb, Cd, Tl, Mo, Al, Ca, Fe, K, Mg, Mn, Na, P) were quantified after extraction with fluoric acid (NF 31–147) followed by ICP-AES quantification [NF ISO 22036]. Exchangeable cations were determined using a cobaltihexamine extract (NF 31–130) and the sum of their charges was calculated to estimate their cation exchange capacity (CEC). Al-Fe oxide contents in the soil were determined using the Tamm method (Tamm, 1922). All soil analyses were made at the INRAE central platform (the LAS laboratory), which has the French quality certification (COFRAC) and participates in ring tests for quality control.
Trees
Survival, growth and nutrition of trees studies were based on 4–5 subplots (depending on the plot surface area). Each subplot was a rectangle of a surface area that depended on the plantation density (WBP=1,600 m2; NuM=1,920 m2; COM=2,688 m2) and where all trees were surveyed. Field campaigns were carried out during each dormant season. Tree height was always recorded and stem circumference at breast height (CBH) has been measured since the trees were 7 years-old. Tree nutrition was assessed during the growing season by sampling 10 trees per subplot. For each tree, a twig directly exposed to sunlight was collected using a precision rifle. Then, intact current-year needles were collected and needles from all of the subplot trees were merged to produce a composite sample. These foliage materials were finally dried, ground and analysed for major nutrients (N, P, K, Ca and Mg) and for Mn, an oligo-nutrient that may be limiting for plants of the study region (Trichet et al., 2018). Nutrients were analysed after wet mineralisation (H2SO4+H2O2). N and P were quantified colorimetrically (AutoAnalyser-3-High-Resolution colorimeter, SEAL) while other elements were quantified by flame atomic absorption spectroscopy (SpectrAA-20, VARIAN). All plant analyses were made using standard reference materials for quality control. We also collected data characterising Pinus pinaster in terms of regional mean values (Saur et al., 1992; Augusto et al., 2008; Trichet et al., 2018) and nutritional threshold values (van den Burg, 1985).
Tree biomass was estimated based on allometric relationships for aboveground compartments (Shaiek et al., 2011), stembark (Augusto et al., 2008), coarse roots (Augusto et al., 2015) and fine roots (Achat et al., 2018). Similarly, foliage surface area, leaf area index and light transmittance rate were estimated based on dedicated empirical models (Berbigier & Bonnefond, 1995; Porte et al., 2000).
Spontaneous vegetation and sown N-fixers
Aboveground biomass and composition of the lowest vegetation stratum (hereafter referred to as “understory”) was monitored using annual destructive samplings. Sampling was carried out in mid-July, which corresponds to the peak of the growing period (Bergeret, 1980; Werno, 1984). The understory was studied in the 4–5 subplots dedicated to trees (see above). In each subplot, four sampling squares (1 m2 each) were displayed to be representative of tree ridges and inter-tree furrows, giving 16–20 squares per plot. In each square, all aboveground plant biomass was collected and taken to the laboratory. Then, biomass was sorted by species, dried and weighed. During the initial years of monitoring, subsamples were used to analyse the nutrient content of the understory (chemical methods described above). The main plant species present in the understory were purple moor-grass (Molinia caerulea), gorses (Ulex europaeus and Ulex minor), common broom (Cytisus scoparius), bracken (Pteridium aquilinum) and ericaceous species (mainly Calluna vulgaris and Erica scoparia).
In March 2013, a 15N labelling area (100 m2) was set up in the NuM plot in order to quantify the percentage of nitrogen derived from the atmosphere in the N-fixer biomass (Danso et al., 1993). In practice, a non-fertilising dose (i.e. 7.5 mg-N m-2) of 99%-pure 15NH4Cl was sprayed onto the bare topsoil. In the subsequent growing seasons, current-year foliage was sampled from N-fixer individuals and from non-fixing plants, hereafter referred to as “reference plants”. The latter individuals were chosen out of a few species that were shown to be reliable reference species (Augusto et al., 2005; Delerue et al., 2015). During each field campaign, five pairs of healthy plant individuals (1 N-fixer + 1 reference species) of similar development stage were selected with the objective of having two individuals growing side-by-side. Samples were dried, finely ground with a ball mill, and δ15N was determined with an inductively coupled plasma (ICP) mass spectrophotometer. δ15N values were determined at the SilvaTech analytical platform, which complies with the ISO 9001:2015 quality certification. The percentage of nitrogen derived from the atmosphere (%Ndfa) was calculated (see equation in Cavard et al., 2007) based on the isotopic δ15N signature of: (1) a reference species, (2) the N-fixer species in field conditions, and (3) the N-fixer species in controlled conditions, with no available N.
Litterfall
The fluxes of dead organic matter arriving on the soil surface were monitored during two consecutive years. For this, 24 litter-traps (i.e. 3 plots × 2 subplots × 4 traps) were installed at the beginning of the 2022 growing season. Each litter-trap is a cubic collector (with an opening of 0.5 m2) with four legs above the understory vegetation, implying that only the tree litterfall was surveyed. Litterfall was collected six-times a year, and then dried, sorted (by tree species and by compartment [foliage, woody, other]) and weighed.
Atmospheric deposition in forest stands
The fluxes of elements were monitored for one year under tree canopies. During the early stages of the experiment, the trees were small and so monitoring was initiated from 10 years-old onwards. Stemflow was assumed to be negligible based on the rough nature of Pinus pinaster bark, a property that limits stemflow (Levia & Frost, 2003), and on local studies (Courcoux, 1982; Loustau et al., 1992a, 1992b). Throughfall was monitored using four gutters per plot (197.5×20.0 cm), equipped with a coarse filter (Thimonier, 1998). The spatial distribution of canopies and gaps modifies the throughfall chemistry (Carleton & Kavanagh, 1990; Seiler & Matzner, 1995; Bottcher et al., 1997; Thimonier, 1998; Levia & Frost, 2006), so the gutters were distributed to be representative of tree ridges and inter-tree furrows. Throughfall was collected, weighed, filtered, and stored at 4°C. Every cycle of four consecutive weeks, a composite sample was made for each collector using the four individual samples (i.e. one per week) and taking their volume into account. This handling implied that there were 13 cycles per year (i.e. 13×4=52 weeks). All composite samples were analysed quickly after sampling for pH and for NO3, NH4 and PO4 (Skalar SAN++). Then, samples were frozen (-20°C) and stored until other analyses could be made (dissolved Ntotal and dissolved organic carbon [DOC] by the combustion method; P, K, Ca, Mg, Mn, Al, Fe, S by ICP-AES). All solutions were analysed using procedures that comply with the COFRAC quality certification (see above).
The element content found in the throughfall collectors do not correspond to atmospheric deposition strictly speaking because some chemical exchanges can occur between tree foliage and rain (Carleton & Kavanagh, 1990; McLaughlin et al., 1996). Consequently we estimated canopy exchanges and atmospheric deposition under the forest canopy using the “budget model” method (Draaijers & Erisman, 1995). This approach uses a chemical element that is not exchanged on foliage as a reference, such as sodium, chlorine or sulphur (Reynolds, 1996; Erisman & Draaijers, 2003; Gonzalez-Arias et al., 2006). In this study, we chose sulphur (S) as it has been shown that it is not be affected by Pinus pinaster canopies (Bellot et al., 1995). The budget model method (Draaijers & Erisman, 1995) states that the canopy exchange (CE) of the reference element is nil (CES=0) and that the ratio {under canopy:open area} of the fluxes of atmospheric deposition of a given element (for instance Ca) is the same for all elements (including the reference element). Based on the reference element assumption and the canopy:open ratio, it is then possible to calculate the fluxes due to canopy exchanges (CE>0 being canopy leaching; CE<0 being canopy uptake) or due to atmospheric deposition (for full details, see Draaijers & Erisman, 1995).
Soil solution composition and soil microclimate
Twenty soil pits per plot were dug at the beginning of the monitoring: 10 pits for each of the two peripheral subplots (Figure S4B). In each subplot, two pits were equipped with TDR sensors at different depths (∼10, 20, 45 and 90 cm) to monitor soil temperature, water content and conductivity (reflectometer Campbell Scientific CS-650; measurement every minute). The reliability of all sensors was checked under laboratory conditions before their installation in the field. Physical soil properties (TDR sensors) were averaged by soil depth based on the four replicates installed in each plot (i.e. two replicates per subplot), which was shown to be a sufficient number to be representative (Schmidinger et al., 2024). Once averaged by plot and soil depth, daily mean values and cycle (4-weeks) mean values were calculated.
In the eight other pits of a given subplot, ceramic suction cups (50 cm long, 2 cm in diameter) were inserted horizontally into the soil. Suction cups were installed at systematic depths (∼20, 45 and 90 cm) because, in podzols, the soil solution composition is not related to soil horizon boundaries (Gottlein & Stanjek, 1996). Depression (0.8 bar) was applied daily to collect extractable soil solution (except during prolonged dry periods to avoid damaging the pumps or the pipe network) and the extracted soil solutions were collected weekly. Soil solutions were analysed using the same handling and methods as for throughfall solutions (see above).
Water table and drainage ditches
Each plot is equipped with three piezometers, one in the central area of the plot and two in the peripheral subplots (Figure S4B). The two piezometers in the peripheral areas were equipped to monitor the water table depth and temperature (piezometer Campbell Scientific CS-451) and their values were averaged. The piezometer in the central area was used to collect water samples four times per year (spring, summer, autumn, winter). If the drainage ditches contained water at the date of water table sampling, they were also sampled (Figure S4A) because they are important contributors to surface water in the study region (Vernier et al., 2003).
In practice, the water table was first pumped during at least 10 minutes and until the properties (pH, O2 concentration, conductivity, temperature) of the water were stable. For this, a multi-parameter sensor (Hanna Instruments, ref. HI 98194; systematically calibrated before fieldwork) was immerged in the recipient where the pumped outflow was directed. Once stability of the water properties was reached, the physical properties were recorded and the odour and colour of the water was described. In addition, water samples were collected, filtered in the field, and some of them were immediately acidified to ensure chemical stability before analysis (DOC, PO43-, Fe2+). Non-acidified samples were analysed for DON, DOP, mineral N, and alkalinity. Acidified water samples and samples for alkalinity were stored at 4°C before analyses. Samples for N compounds were frozen at -20°C. Fe2+, PO43- and dissolved N compounds were measured with colorimetric methods. DON and DOP were measured after mineralisation with persulphate. DOC concentrations were determined by high temperature catalytic oxidation method using a Shimadzu TOC 5000 analyser. All solution analyses were made using standard reference materials for quality control.
Open area monitoring
The station dedicated to monitor the weather conditions and the atmospheric bulk deposition was not located in the study plots but in an open area (44.74185°N, 0.78172°W; 0.5–0.8 km distant from the forest plots) of the INRAE experimental domain. The station is far from any forest edge, tree hedge or building. The weather data (air temperature, precipitation) were collected using the INRAE Climatik webservice (https://agroclim.inrae.fr/climatik). In addition, bulk precipitation was collected using five collectors (170 mm in diameter) each linked to a large, chemically inert, can. All the cans were placed in a box, buried to limit temperature fluctuations. Finally, wet deposition was studied using an automatic precipitation sampler NSA-181 (www.eigenbrodt.de/en/products/sampler-collectors-instruments/nsa-181-basic). All the precipitation volumes were measured weekly and then subsampled for chemical analyses (using the same methods as the throughfall solutions; see above). Occult deposition (e.g. fogs) was considered as negligible (Erisman et al., 1997).
Results and Discussion
Initial soil properties
The soils of the large plots are recognised as podzols (FAO/IUSS & FAO, 2006), which is the common case for the study region (Jolivet et al., 2007). These podzols were described as entic (i.e. no E horizon, with an A/Bs/C profile) in 80% of cases, and ortsteinic (i.e. cemented spodic Bs horizon) in 37% of cases. The combination of these two characteristics led to the following mean distribution, based on the WRB soil classification (FAO/IUSS & FAO, 2006): 14% haplic podzols (A/E/Bs/C), 6% ortsteinic podzols (A/E/Bs-cemented/C), 49% entic podzols (A/Bs/C) and 31% entic ortsteinic podzols (A/Bs-cemented /C). This distribution was similar among the three plots (even if the proportion of entic podzols was slightly lower in the NuM plot (72%)), and was representative of the study region (Augusto et al., 2010) including the spatial variability within a given plot (Duchaufour, 1949; Arrouays et al., 2002; Payet, 2004). The thickness –and the depth– of the soil horizons were also comparable for the three plots: The A (black; e.g. 7.5 YR 3/1 using the Munsell colour system) horizon was 39-47 cm thick (range of mean values), the E horizon (grey; e.g. 10 YR 7/2) was 11-15 cm thick (when present), the Bs horizon (brown; e.g. 7.5 YR 4/4) was 27-37 cm thick (top of the horizon at 40-50 cm deep), and the C horizon (yellow; e.g. 10 YR 7/4) appeared at depths of 73-77 cm. The maximum rooting depth was also similar among the studied plots (69-84 cm). Finally, the thickness of the ploughed layer (due to soil preparation before plantation) was similar among forest management alternatives (FMA; 26-31 cm).
As for the pedological description of the soil profiles, the soil physical-properties displayed fairly comparable conditions between plots. For the soil properties that were analysed with several replicates, the vertical distributions were the same for the three plots (Figure S5) and representative of mesophylous–wet moorland soils of the study region (Augusto et al., 2010). The same pattern was observed for all the studied soil properties, such as particle size distribution, Si-Al-Fe oxides, P content, CEC and exchangeable cations, and elemental composition (Figures S6 and S7). Overall, except for the vertical distribution of Zn (Figure S7K), our soil sampling suggested that the initial soil properties of the three large plots were fairly similar. Although initial homogeneity of soils is not sufficient to claim that further differences are due to experimental treatments (Hurlbert, 1984), it is a necessary prerequisite and we assumed that the further differences could be the consequences of the experimental managements.
Biophysical effects of FMA with different levels of canopy openness: water and temperature
As expected (hypothesis H1), the stand density had an influence on tree LAI, with subsequent consequences on light transmittance and litterfall fluxes (Figure 1). In particular, the densest stand (WBP) showed the highest canopy development and litterfall flux, which is logical since litterfall is positively correlated to stand biomass (Matala et al., 2008). The recorded results were similar to published data about maritime pine in terms of LAI and light transmittance (Loustau & Cochard, 1991; Berbigier & Bonnefond, 1995; Rascher et al., 2011), litterfall dynamics (Courcoux, 1982; Hernandez et al., 1992) and litterfall flux (Courcoux, 1982; Kavvadias et al., 2001).
Similarly to sunlight, the stand density influenced the interception rate of the rainfall (del Campo et al., 2022). The mean values of rain interception (calculated using the annual sums) were: combined objective FMA (COM)=18.6%, nutrient management FMA (NuM)=21.5%, wood biomass production FMA (WBP)=23.6%, and were in line with previous studies about maritime pine forests (Butcher, 1977; Courcoux, 1982; Farrington & Bartle, 1991; Loustau et al., 1992b). It is noticeable that the interception rate in the open stand (COM) was logically higher under the tree ridges (26.8%) than under the furrows (10.3%), and that the proportion of rain reaching the understory strongly decreased when the precipitation intensity was low (Figure S8).
The interception differences in sunlight and rain between FMA may have influenced the soil microclimate. Indeed, soil temperature in the dense stand (WBP) was lower than in other stands (Figure 2), particularly during the 2016-2019 period when the WBP stand reached the canopy closure stage before the other stands. While such a canopy-effect on soil temperature was expected (Yan et al., 2011; De Frenne et al., 2021) and in line with our initial hypothesis H1, it was surprising to observe that the deep soil layers (Figure 2EF) were impacted as much as the topsoil layer (Figure 2D). The effects of canopy openness on soil water content were less straightforward to interpret than the soil temperature because the densest stand did not display the lowest water content as it might be expected considering the observed rain interception rates (H1; Yan et al., 2011). Conversely, the soil of the WBP stand seemed wetter in winter than the COM soil (down to 45 cm deep; Figure 2AB). We interpret this apparent paradox to be the result of several processes. Firstly because of its density, the WRB stand is shady, which induced a progressive decline of the understory (Gaudio et al., 2011). This vegetation stratum contributes up to one third of the forest evapotranspiration (Rivalland et al., 2005; Jarosz et al., 2008), and having a sparse understory may have contributed to wetter conditions in this dense stand. Secondly, a lower evaporation rate from the WBP soil may have been caused by lower temperature values. Thirdly, a denser stand canopy probably limited aerodynamic water vapour transport from the soil surface into the atmosphere. Finally, a thicker forest floor layer in the WRB stand may have limited vapour exchanges at the soil-atmosphere interface. We concluded that, whatever the processes responsible, the stand density appeared to be an important property of young plantations for determining the local microclimatic conditions.
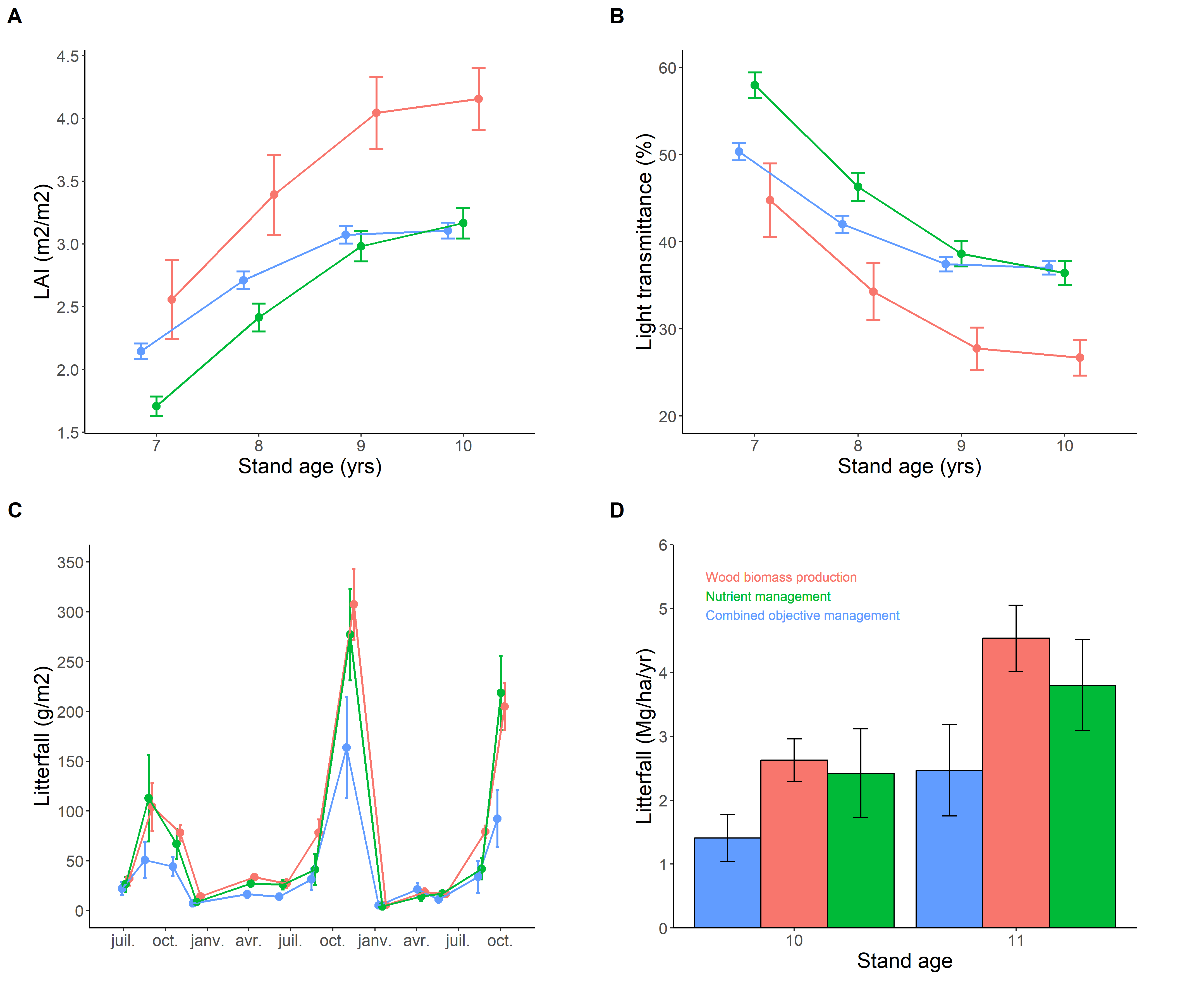
Figure 1 - Stand canopy development and its effects on light transmittance and litterfall. These are mean values ±1 standard error: tree leaf area index (LAI; A), light transmittance through tree canopies (B), litterfall dynamics and annual fluxes (C-D). Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
The FMA had no clear effect on the water table level (Figure 3A). On the other hand, temperature was apparently influenced by the nature of stands, with values that increased with stand openness: COM>NuM>WBP, particularly during the summer (Figure 3B). All in all, from the atmosphere to groundwater, stand density seemed to influence microclimatic conditions. Less water reached the soil in the dense stand, but the soil remained wetter in its upper part. The effects were marked for temperature and were surprising because it was the whole soil-water table continuum that was cooler under the densest stand. This result suggests that silviculture can have important –and literally deep– impacts on local biophysical dynamics.
Nitrogen input through symbiotic fixation of atmospheric N2
In the nutrient management FMA (NuM), the vegetation was left untouched until the sown N-fixers (i.e. common gorse [Ulex europeaeus] and common broom [Cytisus scoparius]) accumulated enough biomass. In practice, while the gorse population developed vigorously, the broom population remained dominated (Figure S9A). This result is in line with the composition of the regional spontaneous vegetation (Barry et al., 1952). Based on the nitrogen content of gorse (Augusto et al., 2009), the N pool size accumulated in total gorse biomass was estimated as 241 kg-N ha-1, which is consistent with other studies about common gorse (Egunjobi, 1971a, 1971b; Augusto et al., 2005). Similarly, based on our 15N labelling, our estimates of the nitrogen fixation rate (i.e. ∼90%) were well in line with current knowledge about this shrub species (Augusto et al., 2005; Cavard et al., 2007; Drake, 2011). Such a high fixation rate implied that the total amount of fixed N was estimated as 217 kg-N ha-1 (i.e. ∼54 kg‑N ha-1 yr-1), which was also in the same order of magnitude as other pine-leguminous associations (Smethurst et al., 1986) and previous studies about pine-gorse associations (Augusto et al., 2005). After four growing seasons, the vegetation was crushed in the NuM plot. However, because only the stand furrows could be crushed (∼75% of the stand surface area; Figure S9B), and because the gorse roots remained alive (enabling further regrowth by sprouting; Figure S9A), we considered that only ∼140 kg-N ha-1 were applied onto the soil as fresh plant debris.
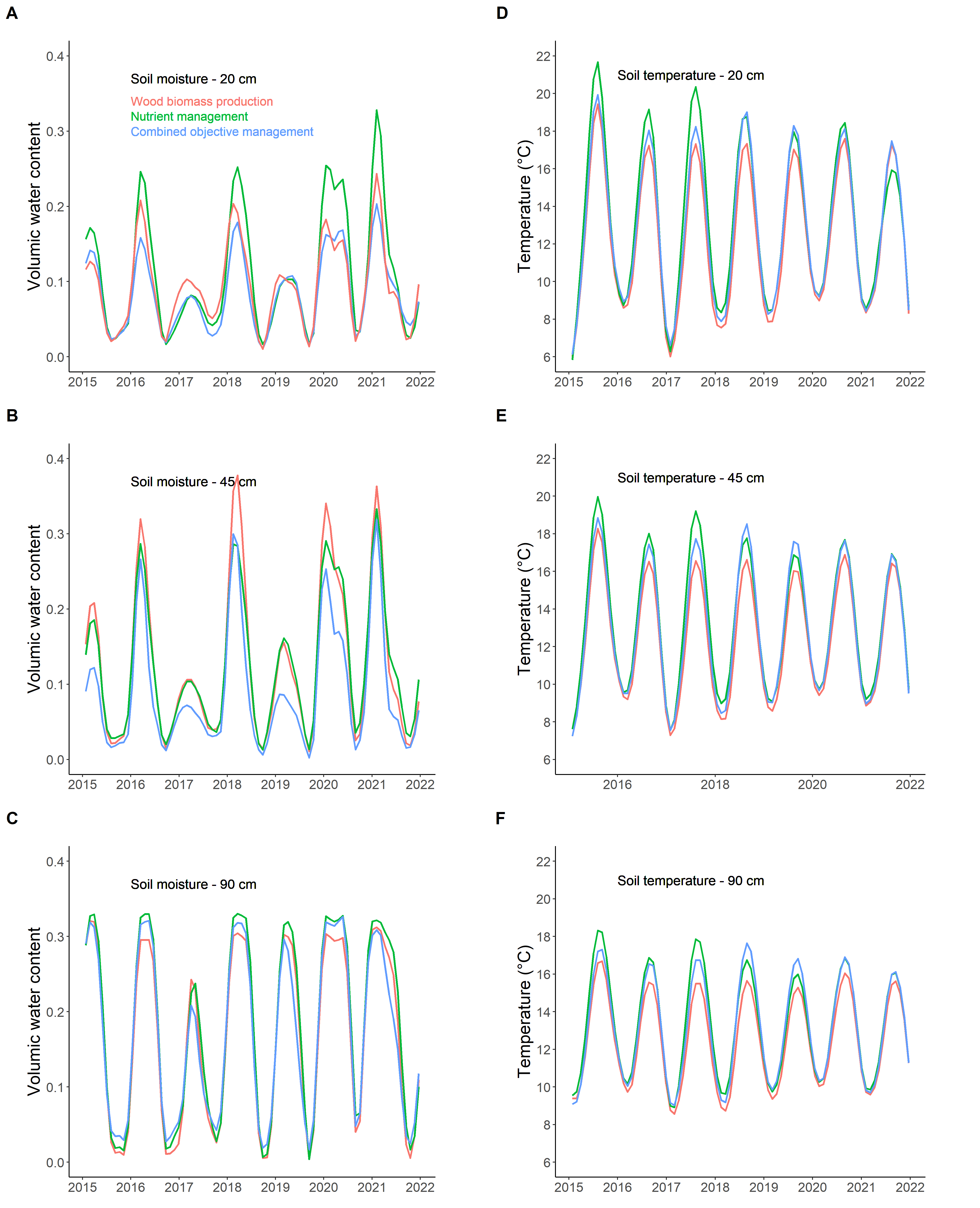
Figure 2 - Soil microclimate. Soil water content (A-C) and soil temperature (D-F) at depths of 20 cm (A,D), 45 cm (B,E) and 90 cm (C,F). Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
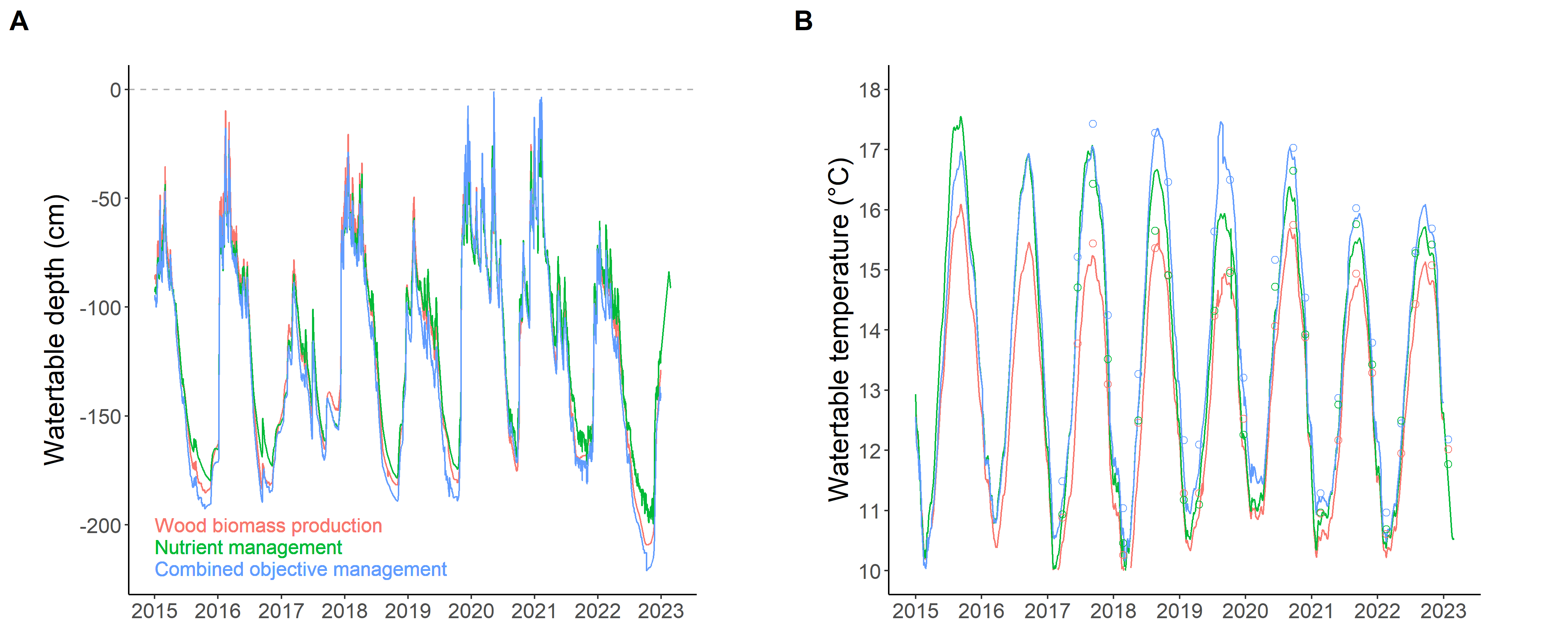
Figure 3 - Watertable level and temperature. Lines represent the mean value (n=2 piezometers) of watertable level (A) and temperature (B). In addition, the temperature values recorded during occasional sampling for water chemistry (n=1 piezometer, distinct from the two other piezometers; see Figure S3B) are shown using dots. Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
Atmospheric deposition of elements and interactions with tree canopies
Atmospheric deposition with no interactions with tree canopies: open area station
The composition of bulk precipitation showed strong relationships between several elements or molecules. Notably, Na, Cl and S were positively correlated, which is probably the consequence of oceanic spray (Gonzalez-Arias et al., 2006; Silva et al., 2007; Vet et al., 2014). Similarly, NO3, NH4 and S were positively correlated with each other, suggesting a pollution origin for N forms and a dual origin (i.e. ocean and pollution) for S (de Vries et al., 2003).
Table 2 - Annual nutrient fluxes due to atmospheric deposition in the open area station. Annual flux values are the mean values of 9 monitored years (mean±1 standard error). The location of the open area station is shown in Figure S1C.
Nutrient | Bulk precipitation | Wet precipitation | Dry precipitation | %Dry | Wet:Bulk ratio |
|---|---|---|---|---|---|
- | (kg ha-1 yr-1) | (kg ha-1 yr-1) | (kg ha-1 yr-1) | (%) | slope (r2) |
NO3- | 4.48 ± 0.82 | 4.82 ± 0.62 | 0 ± 0# | 0%# | +0.75 (0.72) |
NH4+ | 6.63 ± 1.60 | 2.66 ± 0.16 | 4.08 ± 1.51 | 60% | ¤ (0.25) |
N-mineral | 11.11 ± 1.92 | 7.47 ± 0.75 | 4.06 ± 1.85 | 33% | ¤ (0.26) |
P | 0.176 ± 0.031 | 0.105 ± 0.021 | 0.076 ± 0.029 | 40% | ¤ (0.34) |
K | 2.42 ± 0.35 | 1.21 ± 0.27 | 1.21 ± 0.14 | 50% | +0.57 (0.82) |
Ca | 6.47 ± 0.62 | 2.96 ± 0.50 | 3.52 ± 0.34 | 54% | +0.57 (0.83) |
Mg | 4.17 ± 1.01 | 2.90 ± 0.68 | 1.28 ± 0.34 | 30% | +0.76 (0.96) |
S | 4.69 ± 0.87 | 3.13 ± 0.51 | 1.56 ± 0.40 | 33% | +0.69 (0.96) |
Cl | 44.21 ± 4.47 | 32.94 ± 3.64 | 11.27 ± 1.77 | 25% | +0.80 (0.95) |
(#) The negative value was corrected as a nil value. (¤) only slope values of regressions with r2>0.50 are presented. N-mineral was calculated as the sum of NO3- and NH4+. Bulk precipitation=wet precipitation+dry precipitation
Linked to the proximity of the ocean (Courcoux, 1982; Amezaga et al., 1997; Silva et al., 2007), the annual fluxes in the bulk deposition were dominated by oceanic NaCl salts (Table 2). The other nutrient fluxes ranged between 2.4 and 6.5 kg ha-1 yr-1, with the notable exception of P (<0.2 kg ha-1 yr-1). These fluxes were consistent with the literature (Courcoux, 1982; Campbell et al., 2004; Croise et al., 2005; Watmough et al., 2005; Vet et al., 2014), including the very low deposition rate of P (Vicars et al., 2010; Jarosiewicz & Witek, 2016; Sohrt et al., 2019). Except for NO3 inputs that were almost completely derived from wet deposition, other inputs arrived both as wet deposition and dry deposition (Table 2). The relative proportions of wet versus dry deposition varied according to the element and were consistent with published results (Erisman et al., 1997; Balestrini et al., 2007). Similarly, deposition fluxes as sampled by a wet-only collector (i.e. wet deposition) was systematically lower than fluxes estimated based on open collectors (i.e. bulk deposition; Table 2), with values that were fairly consistent with the literature (Lindberg et al., 1986; Staelens et al., 2005; Balestrini et al., 2007).
Atmospheric deposition with interactions with tree canopies: forest stands
The FMA had no effect on pH mean values of throughfall (i.e. pH=4.7-4.8). Similarly, forest management had no clear influence on the chemical composition of the throughfall, which was collected below tree canopies (Table S3). This was in line with our initial expectation (H2). The chemical properties of the throughfall in our study were fairly similar to those of previous studies about maritime pine south-western France (Table S3; Pitaud, 1967; Courcoux, 1982).
The ratio between throughfall and open deposition was close to 1 for elements not exchanged on the foliage (i.e. Na and S), with no effect of FMA. This negligible amplifying effect of tree canopies on atmospheric deposition was not surprising (hypothesis H2) because trees were still young and small, and because tree height is a major driver of forest efficiency to capture atmospheric deposition (Erisman & Draaijers, 2003). Such a pattern has already been reported for a young Pinus pinaster stand (Courcoux, 1982). Based on this result, we estimated canopy exchange assuming that it was the difference between throughfall and bulk deposition, and we found no clear effect of FMA on throughfall and canopy exchange. In general, throughfall was enriched in potassium and, to a lower extent, in magnesium due to canopy leaching (Figure 4). In practice, 62-66% of K and 7-13% of Mg in the throughfall originated from foliage leaching, a pattern that is consistent with many field studies (Morris et al., 2003; Tomaszewski et al., 2003; Chiwa et al., 2004). Conversely, mineral nitrogen (NO3, NH4) contained in bulk deposition was partly absorbed by tree foliage before reaching the soil surface (26-34% of NO3; 67-72% NH4; 39-48% N-mineral; Figure 4), also in line with published studies (Courcoux, 1982; Edmonds et al., 1991; Morris et al., 2003; Chiwa et al., 2004).
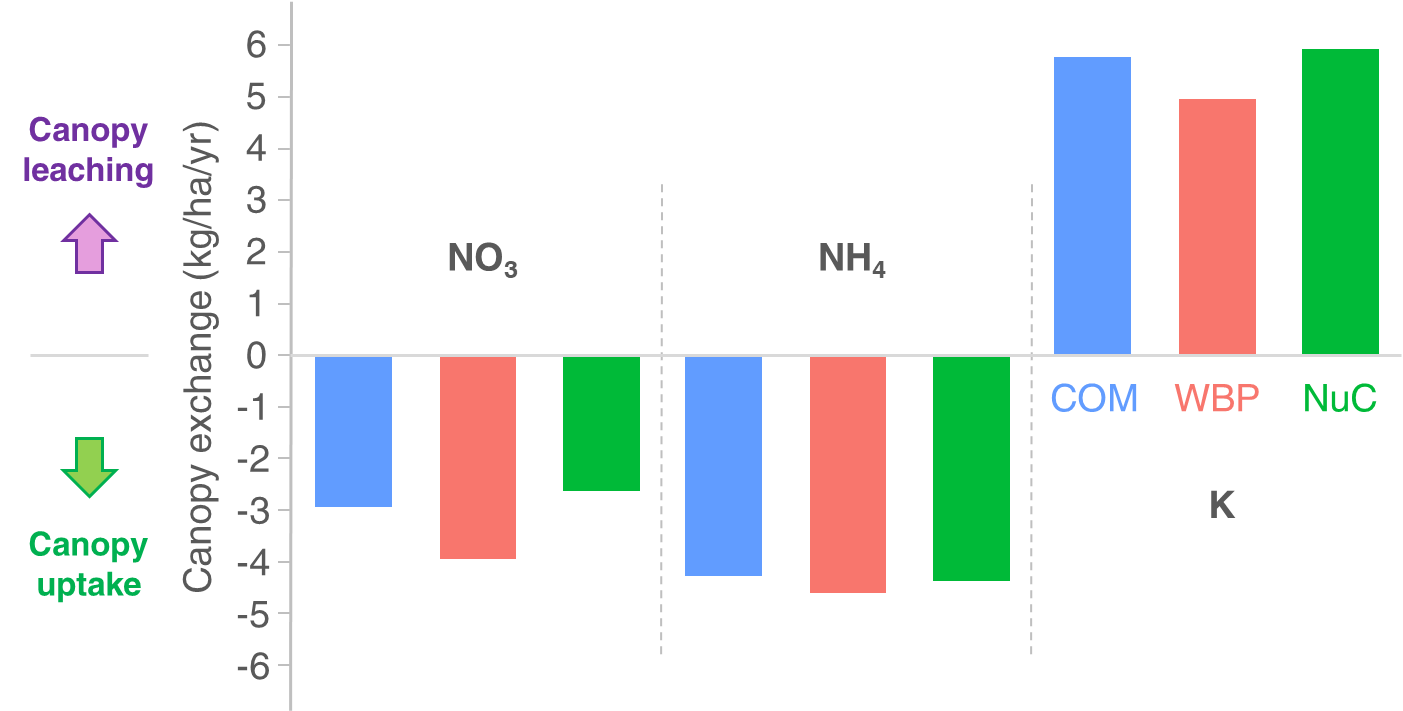
Figure 4 - Chemical canopy exchanges. Annual fluxes of canopy exchanges for nitrogen (in NO3 or NH4 forms) and potassium (K) during the passage of rain on the foliage of the trees. Positive values indicate nutrient leaching from pine needles to throughfall, whereas negative values indicate uptake of nutrients by trees from rainwater. See Methods for more details about calculations. Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
Soil solution biogeochemistry
The soil solution chemistry was characterised by high concentrations of dissolved organic matter, including carbon (DOC) and nitrogen (DON). These compounds showed a logical decrease with increasing soil depth (Figure S10) and seasonal variations (Kaiser et al., 2003). Such a pattern is commonly reported for ecosystems with perennial vegetation (Danielson et al., 2017) and was consequently expected in this forest context (Mignot-Delprat, 1997). The nutrient management (NuM) FMA showed particularly high concentrations of dissolved organic matter in the topsoil layer (Figure S10A,D). The prominence of organic forms is typical of undisturbed ecosystems (Qualls, 2000; Danielson et al., 2017), but this general trend was not observed in the topsoils of the combined-objective management (COM) and wood biomass production (WBP) FMA as dissolved nitrogen was dominated by NO3 (Figure 5A,D). This difference between FMA was not the consequence of differences in NO3, which showed similar ranges of values for the three FMA, but was the result of the high concentration values in DON in the NuM FMA. This pattern was surprising because the NuM understory was dominated by N-fixers (mainly Ulex europaeus) that are known to increase N-mineralisation (Smethurst et al., 1986; Rozé, 1987; Mo et al., 2016) and hence the nitrate concentration of soil solutions (Dyck et al., 1983; Magesan et al., 2012; Vidal et al., 2019). This unexpected trend, which partly conflicted with our hypothesis H3, may be explained by the huge differences of understory biomass between FMA. Indeed, while the aboveground biomass peaked in the first years at 5.8 Mg ha-1 (COM) and 4.6 Mg ha-1 (WBP), and then decreased as a result of vegetation control (COM) or tree canopy development (WBP), it had continued to increase in the NuM FMA (Figure S9A). Such a high level of standing biomass produced high amounts of dead organic matter after understory crushing (Figure S11) that had partly dissolved in the soil solution (Chapman et al., 2001). A sharp increase of the proportion of dissolved N as DON (Figure 5A,D) due to the application of large amounts of fresh woody debris is consistent with the literature (Chang et al., 2007; Morris, 2009) and probably resulted from the high N-demand of the microbes that were decomposing the applied debris (Berg, 1988; van Wesemael, 1993; Moro & Domingo, 2000). This pattern is also consistent with the timing of the experiment because the differences between FMA appeared in 2017 (Figure 5A,D), which was concomitant with the crushing of the NuM understory (Figure S9). This difference of soil solution composition probably also explains the more acidic conditions observed in the NuM soil (Figure 5G), as dissolved organic matter in this kind of podzols is known to contain various organic acids (Juste, 1965; Mignot-Delprat, 1997). Conversely, the presence of an abundant layer of living N-fixers did not induce nitrate leaching in the deep soil layers, as it may have been anticipated based on the literature (H3; Drake, 2011).
Another possible difference between FMA regarding soil solutions was their ionic strength as estimated by electrical conductivity (Simón & Garcı́a, 1999). For this property, the FMA seemed to rank as follows: WBP>NuM>COM (Figure S12A-B). Soil solution chemistry is often dominated by calcium (Schneider, 1997), even in acidic forest soils (Augusto & Ranger, 2001) such as these podzols (Merzeau, 1983), and so we interpreted the high conductivity values under the WBP stand as the consequence of high Ca concentrations (Figure S12D-E), but also as a result of different soil water moisture levels (Figure 2A-B). Nevertheless, because of the observational nature of the present study, this possible difference needs further investigation for confirmation.
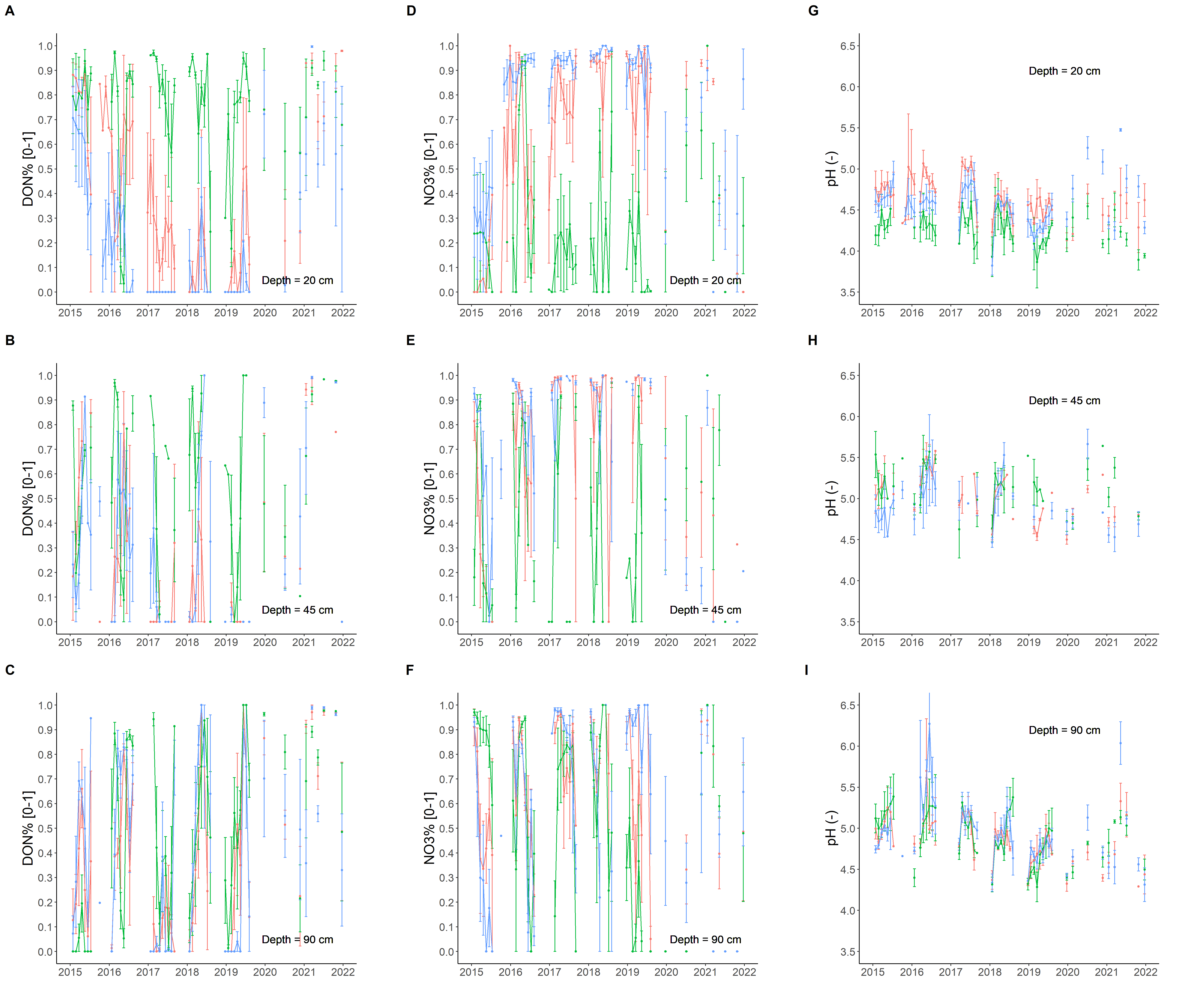
Figure 5 - Soil solution nitrogen. DON%: percentage of total nitrogen composed of dissolved organic nitrogen (A-C), NO3%: percentage of total nitrogen composed of NO3 (D-F), pH value (G-I). Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
Possible biogeochemical consequences on the water table and drained waters
The water table was characterised by acidic conditions, with large fluctuations between pH=4 and pH=5, but without any clear influence of FMA (Figure 6A) except water conductivity that seemed to be slightly lower under the COM as compared with other FMA. The striking result in the conductivity dynamics was not related to FMA but to the large peaks in 2020-2021, 2023 and, to a lesser extent, 2018 (Figure 6B). These peaks generally coincided with periods of high water table levels (Figure 3). The water table was also characterised by low values, and erratic variations, in dissolved oxygen (Deirmendjian et al., 2019), with no clear seasonal pattern (Figure 6C). These variations were negatively associated with Fe2+ concentrations (r=-0.36; P=0.001; Deirmendjian et al., 2019), and turbid and smelly (H2S odour) water (χ2=10.7, P=0.014), which were collected mainly during the winter season. Finally, the water table composition was characterised by low nutrient concentrations such as nitrogen, which was often below 1 mg L-1 and always below 2 mg L-1 (Figure 6D).
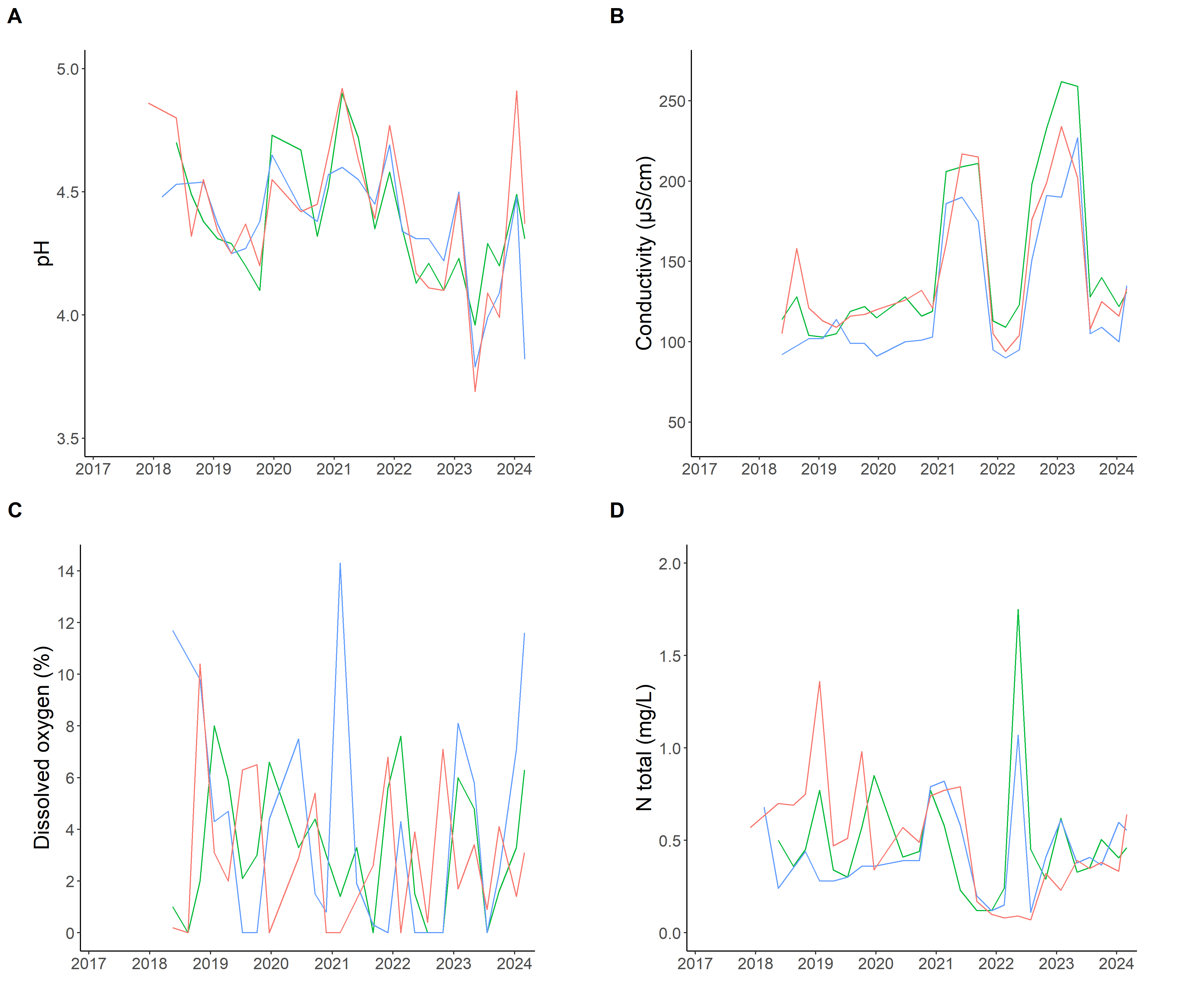
Figure 6 - Watertable physico-chemical properties. Red = wood biomass production (WBP); green = nutrient management (NuM); blue = combined objective management (COM).
In drainage ditches, the water chemistry showed a negative relationship between DOC and dissolved oxygen (DO2; r=-0.38; P=0.013). More precisely, during the winter, the DOC values increased while the DO2 values decreased along the drainage route (Figure S13A-B), indicating concomitant re-oxygenation and mineralisation (Deirmendjian et al., 2018). In line with the water table dynamics, we observed that the electrical conductivity of the water in drainage ditches doubled during the periods of high water level as compared with periods of low water level (Figure S13C). It is probable that high water levels dissolved large amounts of organic matter located in the topsoil layers, which were then found in the water table and in ditches (Inamdar & Mitchell, 2007; Deirmendjian et al., 2018, 2019). Nitrate concentrations in ditch waters were generally low, but some high values were observed during several winters and were at the expense of DON values (Figure S13D). These periods of high NO3 values were however uncommon as water chemistry was generally dominated by organic forms (Hedin et al., 1995; McGroddy et al., 2008). Whatever the chemical form of nitrogen, drainage ditches were probably a major vector of nutrient outputs as they were identified as the main contributors to the surface waters of the study region (Rimmelin, 1998; Vernier et al., 2003; Deirmendjian et al., 2018).
Although it was not possible to quantify nutrient leaching in a reliable way, our results suggested that nutrients losses though water outputs were small. Indeed, the concentrations of soil solutions at 45 cm and 90 cm remained low compared with other forest ecosystems (Yanai, 1991; Attiwill & Adams, 1993; Berden et al., 1997; Ranger et al., 2001, 2007; Jussy et al., 2004; van der Salm et al., 2007; Sohrt et al., 2019). In addition, the water table chemical composition showed no obvious symptom of pollution and was more oligotrophic than the mean values of the study region (Courcoux, 1982; Merzeau, 1983; Jolivet et al., 2007). Our data interpretation is also consistent with the literature, which showed that N leaching in forests is related to atmospheric N deposition and forest floor C/N ratio (van der Salm et al., 2007). Notably, N leaching increases with increasing atmospheric N deposition (Templer et al., 2022), and is generally negligible when atmospheric deposition is lower than 14 kg-N ha-1 yr-1 (van der Salm et al., 2007), which was the case of our experimental platform (Table 2). Similarly, N leaching in forest ecosystems is generally negligible when the forest floor C/N ratio has values higher than 30 (Gundersen et al., 1998), which is the case of Pinus pinaster stands (≈34-44; Augusto et al., 2010). Finally, our data interpretation is consistent with other studies that were carried out in the same region, which all concluded that N leaching and P leaching were lower than respectively 2.0 kg-N ha-1 yr-1 and 0.2 kg-P ha-1 yr-1, mainly as organic forms (Courcoux, 1982; Campagne, 1997; Rimmelin, 1998; Vernier et al., 2003; De Wit et al., 2005). To sum up, we concluded that nutrient losses through soil leaching were probably of low magnitude. Nevertheless, further investigations are still needed, because simulations produced by the GO+ model (Moreaux et al., 2020) indicated that the annual water seepage from soils was substantial (500±66 mm yr-1) and even more important during years with high rainfall (total range over a decade=221-808 mm yr-1; r=+0.90 between annual rainfall and annual seepage).
Vegetation response to forest management alternatives
Tree nutrition in the study region is characterised by deficiencies in several nutrients, notably potassium and magnesium (Demounem, 1979) and above all phosphorus (Trichet et al., 2009), which explains the common use of P fertilisation in local silviculture (Table 1). Fertilisation at plantation probably improved tree nutrition as shown by foliage P content (Figure 7B), but foliage P content decreased over time and the same trend was visible for most nutrients (Figure 7A-E), suggesting a phenomenon of nutrient dilution induced by tree biomass growth (Jarrell & Beverly, 1981; Manghabati et al., 2019). This decreasing trend was broken for N and K after five years, which coincided with the first major operations of understory control (Figure S9), as the N-K foliage content increased sharply and remained at high levels (Figure 7A,C). Spontaneous vegetation may be competitors of trees (South et al., 2006) and the composition of the local vegetation (Pteridium aquilinum, Molinia caerulea, Rubus fruticosus, Calluna vulgaris and Ulex europaeus) is well-known at the national scale (Dumas et al., 2022) and at the regional scale (Vidal et al., 2021; Bon et al., 2023) for being particularly competitive species. Thus, reducing the biomass of the spontaneous vegetation probably reduced the competition for nutrient uptake. In addition, vegetation crushing produced large amounts of fresh litter that probably released nutrients quickly, such as potassium (Bessaad & Korboulewsky, 2020) and nitrogen (Vidal et al., 2019). The foliage nutrient content showed no difference between FMA, except for a lower P content for NuM and a lower K content for WBP after the first vegetation control. Nitrogen nutrition was not improved by the introduction of N-fixers in the NuM stand, in contradiction to our a priori expectation (H3). These trends will be further investigated using the trial site (in completely randomised blocks) of the XyloSylve platform.
Contrary to foliage nutrient data that showed no –or only minor– differences between FMA, the tree growth data clearly differentiated the three tested silvicultural treatments. One important difference between FMA was the initial stand density (Table 1). As expected, because high stand density induces high competition among trees (Lee & Choi, 2019), we observed that tree survival rate (Figure 8A) and tree growth (Figure 8B-C) were high where the stand density was low. In addition, trees invested relatively more in aboveground biomass with increasing stand density (WBP FMA; Figure 8D), probably as a consequence of competition for access to sunlight (Ilomaki et al., 2003). At the stand scale, however, the WBP FMA had the highest stand biomass, because having small individuals was compensated by having numerous individuals (Figure 8E-F). After 10 growing seasons, the WBP FMA accumulated 33% and 22% more biomass than COM and NuM, respectively. In other words, the most intensive FMA produced the largest amount of biomass, which explains why it was also the WBP FMA that modified the biophysical functioning of the ecosystem the most.
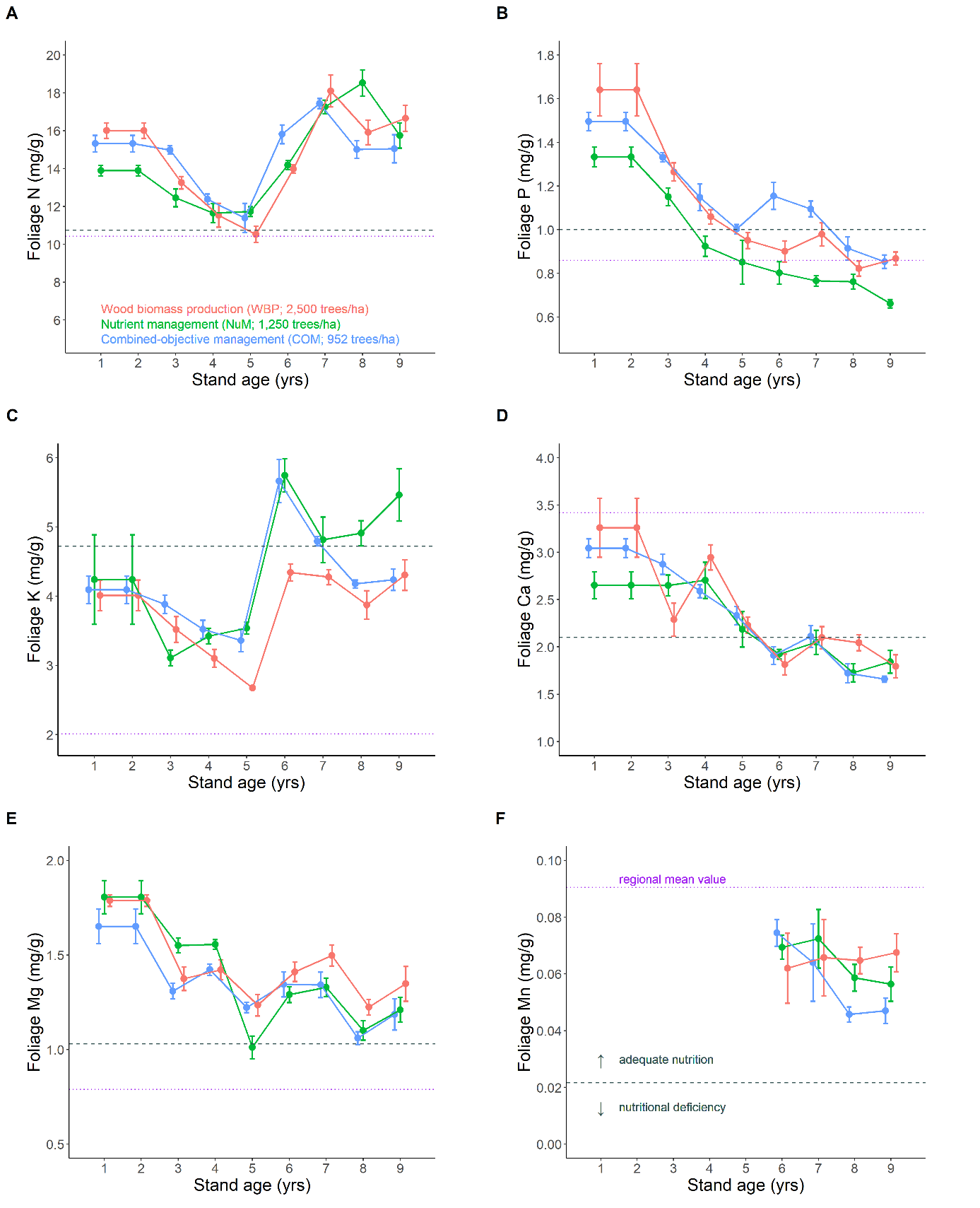
Figure 7 - Tree nutrition. Foliage (1-year-old needles) nutrient content over time. Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM). Dashed grey lines indicate nutritional threshold values between adequate nutrition and deficiency for Pinus pinaster (see F). Dotted purple lines indicate the mean value of the study region for the same tree species.
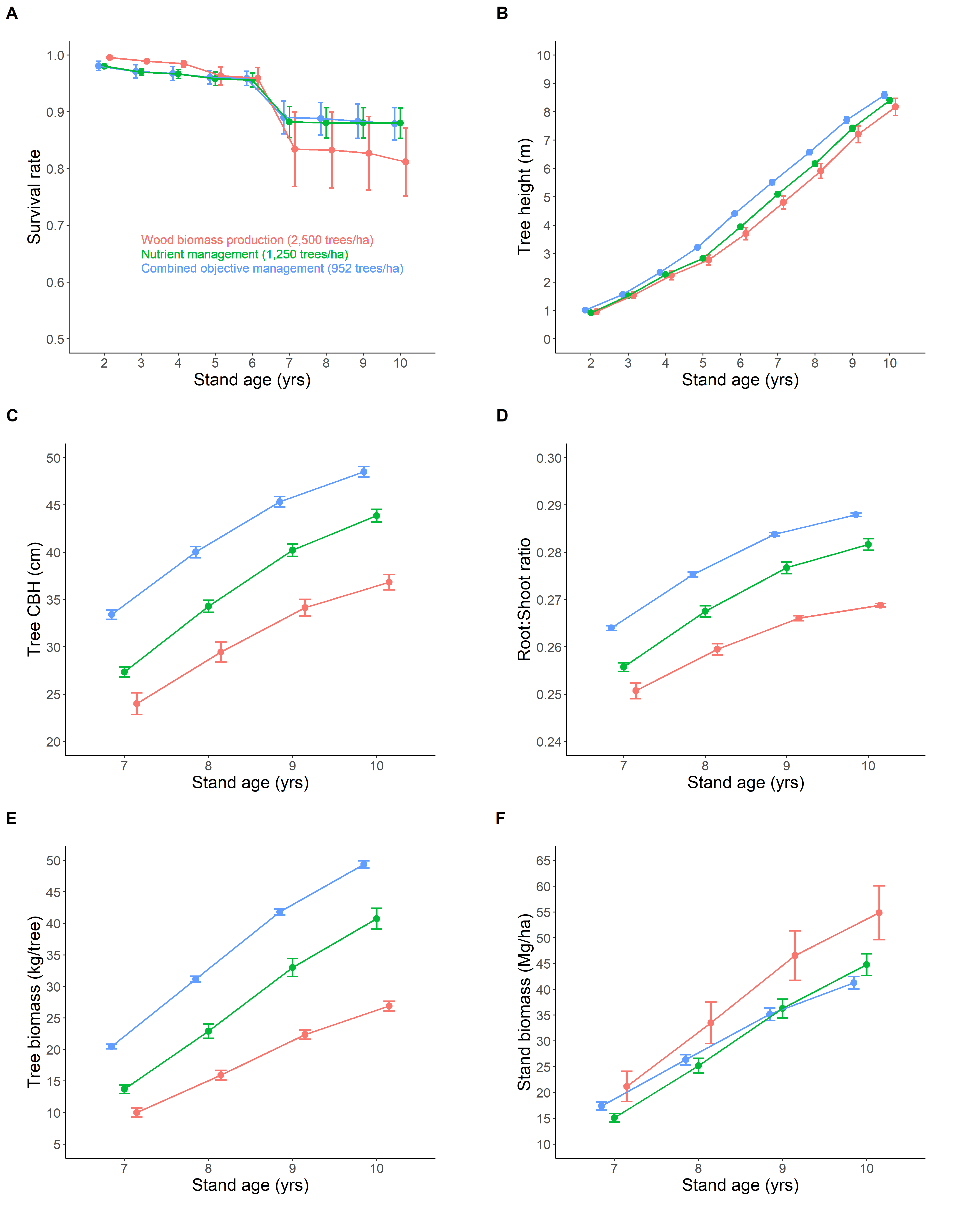
Figure 8 - Tree growth and stand biomass dynamics. Red=wood biomass production (WBP); green=nutrient management (NuM); blue=combined objective management (COM).
Conclusions and perspectives
Our study took place in a field experimental platform characterised by a temperate climate but also by particularly unfavourable edaphic conditions. Soils are acidic sandy podzols and inherently poor in nutrients. In such a context, the nutritional sustainability of forest stands depends on fertiliser inputs (Heilman & Norby, 1998), symbiotic N fixation (Augusto et al., 2005) and atmospheric deposition (De Schrijver et al., 2008). Taking into account the demand for nutrients by pine trees (Helmisaari, 1995; Augusto et al., 2010), it is probable that the current rate of atmospheric deposition is high enough to supply trees with nutrients (except for P, which is supplied through fertilisation), which is well in line with other field studies carried out in unpolluted regions (Campo & Gallardo, 2012; Johnson & Turner, 2014). Although it was not possible to calculate input-output budgets reliably, because of the methodological constraints associated with this approach (Ranger & Turpault, 1999; Paré & Thiffault, 2016), it is probable that budgets are close to equilibrium (or in surplus for N in the NuM FMA) at the stage of stand development. The effects of contrasting FMA on the biogeochemical functioning of ecosystems were modest considering the large structural differences of stands including nitrogen cycling, despite a large symbiotic fixation flux in the NuM FMA. Nonetheless, it is worth mentioning that the trees remained small during a long initial period, probably delaying the biogeochemical consequences of FMA. This is particularly important for the soils studied, because the functioning of podzols is very dependent on soil organic matter (Juste, 1965; Lundstrom, 1993; David et al., 1995; Jansen et al., 2005; Augusto et al., 2010), a compartment of the ecosystem that changes slowly over time (Korner, 2003; Balesdent et al., 2018). Consequently we anticipate that the influence of FMA on ecosystem biogeochemistry will increase with increasing tree age and management modifications. In addition, the tested FMA have designs that include important treatments at mid-stage development. In practice, the NuM FMA will receive a wood ash application to supply the ecosystem with nutrients (Gomez-Rey et al., 2013), while WBP will be thinned and COM will be planted with Mediterranean oak species. The functional consequences of these managements will be assessed in the monitoring site of the XyloSylve platform but also in its trial site, which will enable to statistically test the observed effects.
FMA had stronger effects on biophysical processes (light transmittance, rainfall transmittance, temperature and moisture) and biomass allocation in the ecosystem than on biogeochemical processes. Within the scope of this study, the most intensive FMA –based on dense plantation dedicated to wood biomass production (WBP)– showed important effects on stand microclimate with higher moisture and lower temperature. This density-microclimate interaction in young plantations may be an important indication of how forestry could be adapted to climate change, as the study region is possibly exposed to a future reduction of plant growth due to water stress (Loustau et al., 2005). This is particularly relevant in the driest parts of the Landes de Gascogne forest, which are known to be water-limited, whereas the wettest parts (where the XyloSylve platform is located) are P-limited (Trichet et al., 2008, 2009; Bon et al., 2023).
Finally, although the relationship between stand density and superficial microclimate was expected (Rambo & North, 2009; Kovács et al., 2017), the recurrent effect on deep soil and water table temperature was surprising and to our knowledge not yet reported. Also, taking into account the possible role of the water table for tree water supply (Vincke & Thiry, 2008), we posit that future studies about forest functioning should include the water table in the atmosphere-vegetation-soil continuum.
Acknowledgements
The XyloSylve project was initiated and designed with important contributions from Patrick Pastuszka and Jean-Michel Carnus. We thank Alexandre Bosc, Jean-Paul Chambon, Nicolas Cheval, Laurence Denaix, Matthieu Le Gall, Sylvie Niollet, Luc Puzos, Annie Raffin, Laurent Severin and the BEF-P3F laboratories for their help during the setting-up of the forest stands and the devices dedicated to monitoring. We are particularly grateful to Dominique Gelhaye who kindly provided advice and participated in the fieldwork for installing some of the biogeochemical equipments. The monitoring, data management, samples analyses and platform functioning would not have been possible without the help of the following colleagues, whom we thank: Mark R. Bakker, Lucie Bon, Sylvie Bussière, Cécile Coriou, Bernard Dokhelar, Nathalie Gallegos, Cyriane Garrigou, Frédérique Générale, Sylvie Milin and David Vidal. The monitoring also benefited from collaborations with INRAE’s laboratories for analyses: LAS laboratory (for soils and soil solutions; https://las.hautsdefrance.hub.inrae.fr), USRAVE laboratory (for plants; https://eng-usrave.bordeaux-aquitaine.hub.inrae.fr), and SilvaTech platform (for isotopic analyses; https://silvatech.isc.inrae.fr). Finally, we thank Marie Charru, Céline Meredieu, Aldyth Nys, and Maude Toïgo for their help during the redaction of the present article.
Preprint version 2 of this article has been peer-reviewed and recommended by Peer Community In Forest and Wood Sciences (https://doi.org/10.24072/pci.forestwoodsci.100187; Lockwood, 2025).
Funding
The installation of the experimental platform was made possible by the financial support of the Nouvelle-Aquitaine regional council and the French National Agency for Research (“Agence Nationale de la Recherche” [ANR]; project PIA-1 Xyloforest). The XyloSylve platform functioning was supported by the French ANR, AnaEE-France (www.anaee-france.fr/en), the PEPR FairCarboN (www.pepr-faircarbon.fr/eng) and the GIS-PMF (www.gisgpmf.fr). Jean-Baptiste Ornon was supported by AXA-Climate.
Conflict of interest disclosure
The authors declare that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article. The first author (L.A.) declares being recommender for the Peer Community in Forest and Wood Sciences, but having had no interaction with the board –nor with the reviewers– outside the normal procedure of evaluation.
Data, script, code, and supplementary information availability
Data (CSV files) and scripts (R files) are available at the French Datagouv website (https://doi.org/10.57745/NEBK4J, Augusto, 2025). Supplementary information is available along with the pre-print at the HAL website (https://hal.inrae.fr/hal-05070633, Augusto et al., 2025).